

Formation of cortical fields on a reduced cortical sheet
- PMID: 10559402
- PMCID: PMC6782964
- DOI: 10.1523/JNEUROSCI.19-22-09939.1999
Formation of cortical fields on a reduced cortical sheet
Abstract
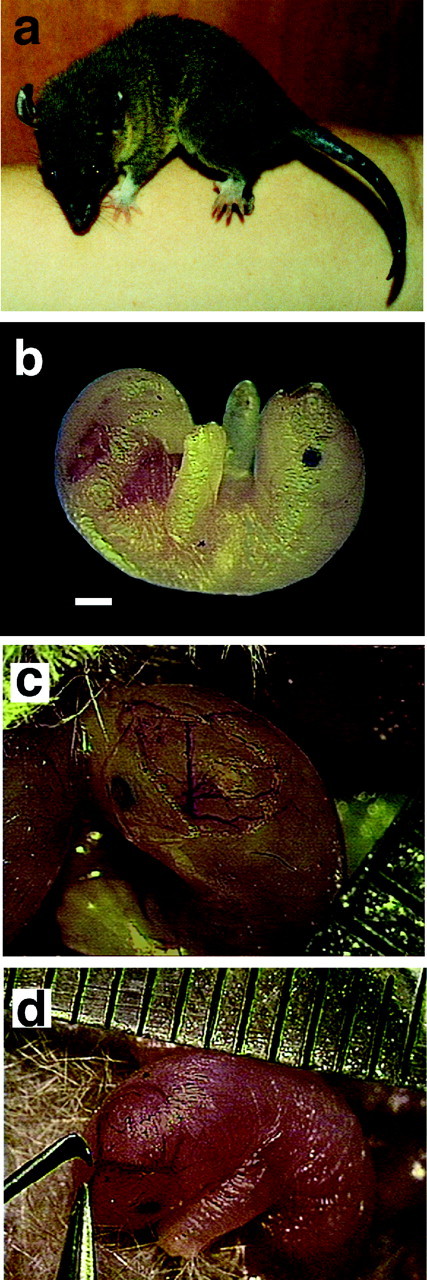
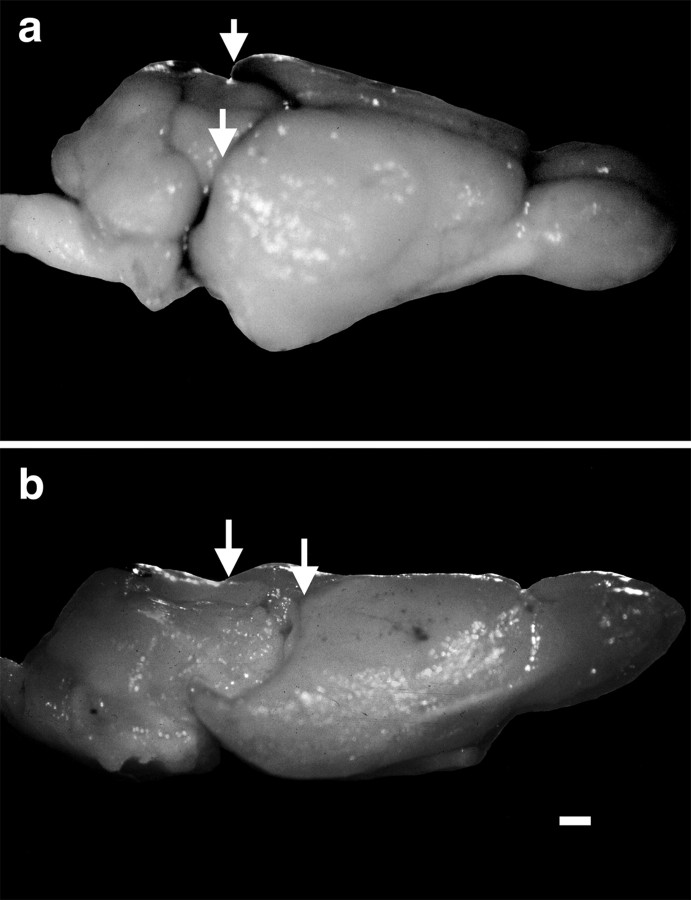
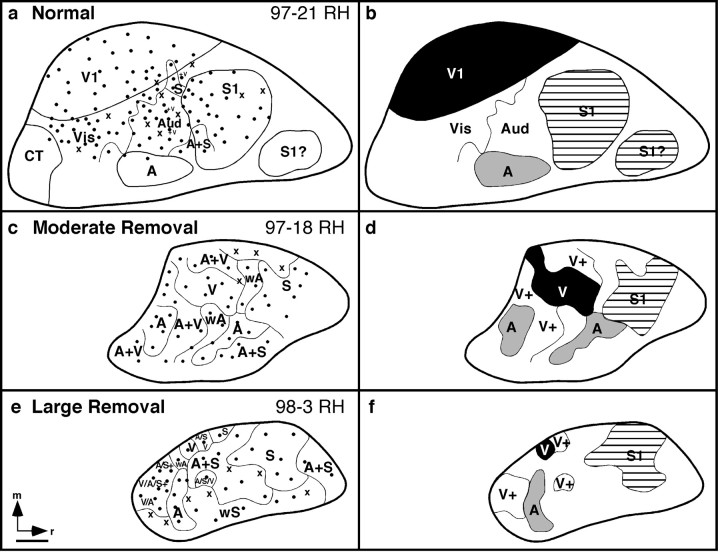
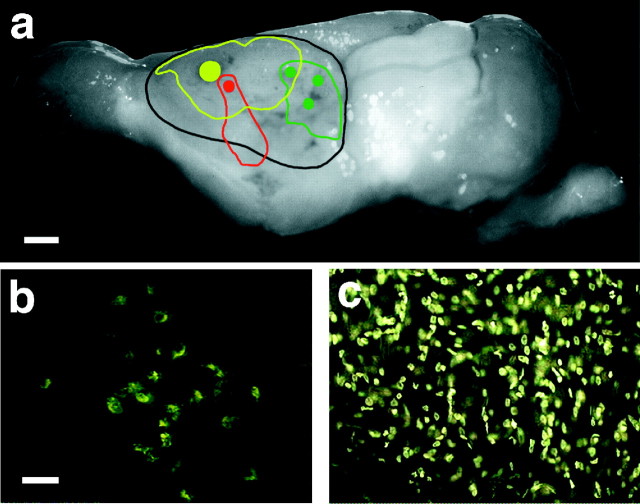
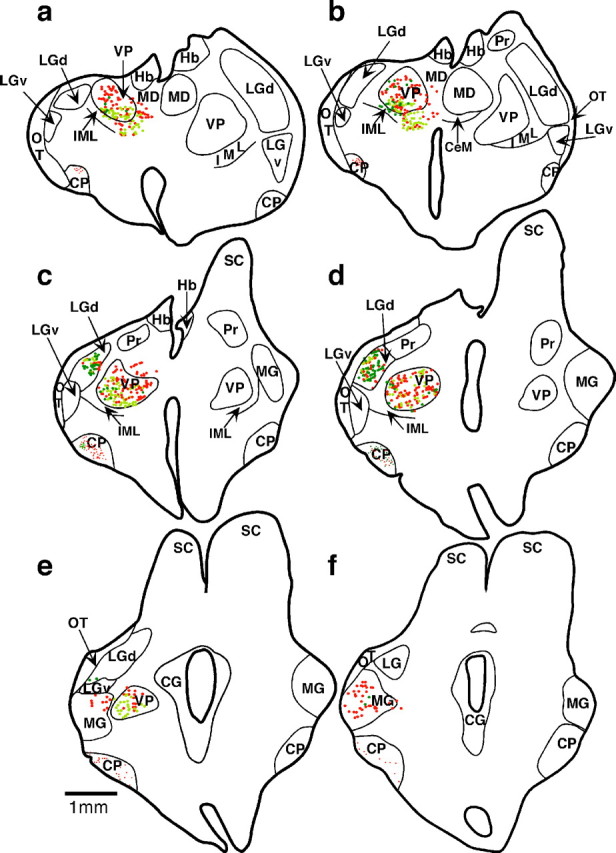
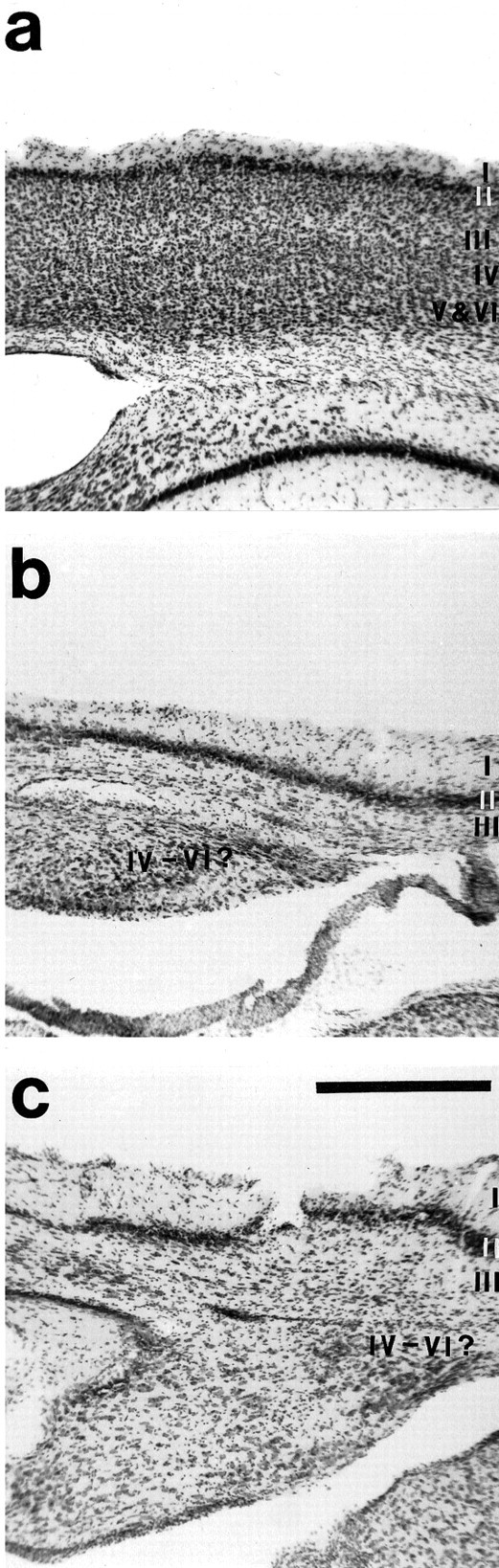
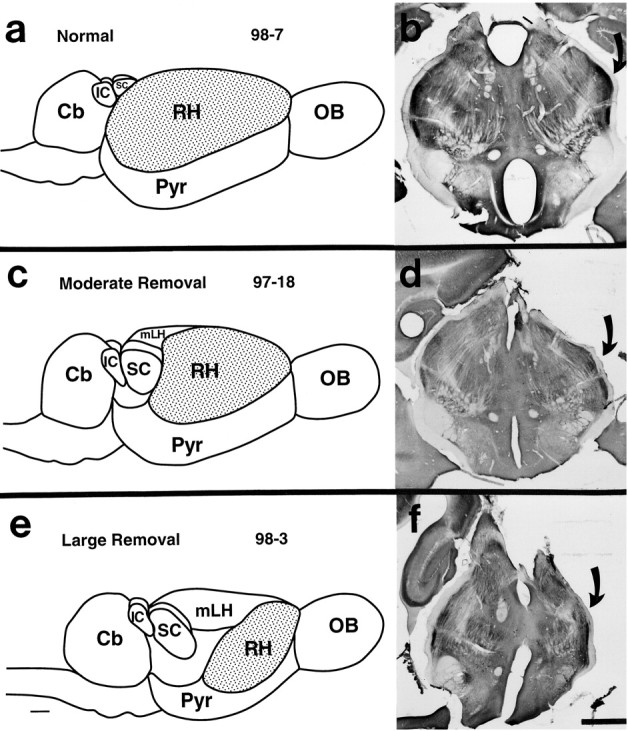
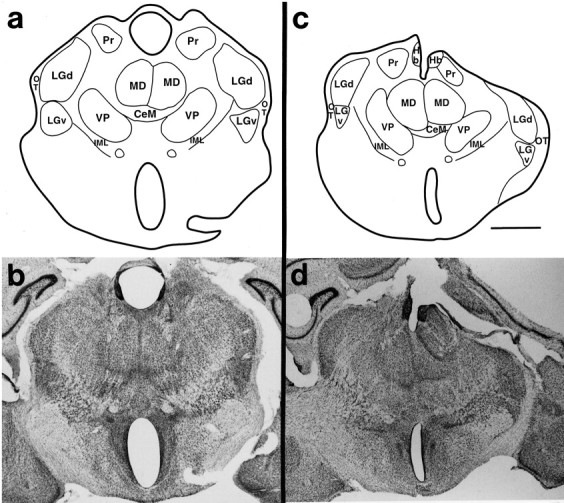
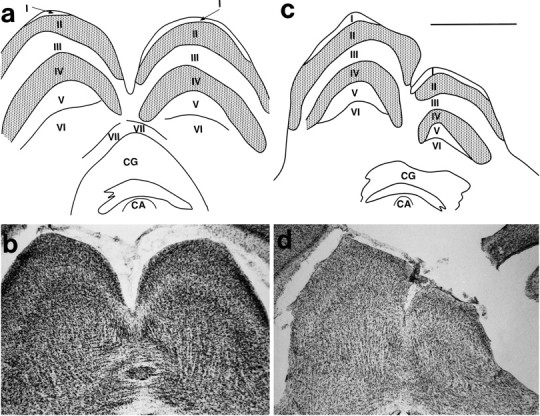
Theories of both cortical field development and cortical evolution propose that thalamocortical projections play a critical role in the differentiation of cortical fields (; ). In the present study, we examined how changing the size of the immature neocortex before the establishment of thalamocortical connections affects the subsequent development and organization of the adult neocortex. This alteration in cortex is consistent with one of the most profound changes made to the mammalian neocortex throughout evolution: cortical size. Removing the caudal one-third to three-fourths of the cortical neuroepithelial sheet unilaterally at an early stage of development in marsupials resulted in normal spatial relationships between visual, somatosensory, and auditory cortical fields on the remaining cortical sheet. Injections of neuroanatomical tracers into the reduced cortex revealed in an altered distribution of thalamocortical axons; this alteration allowed the maintenance of their original anteroposterior distribution. These results demonstrate the capacity of the cortical neuroepithelium to accommodate different cortical fields at early stages of development, although the anteroposterior and mediolateral relationships between cortical fields appear to be invariant. The shifting of afferents and efferents with cortical reduction or expansion at very early stages of development may have occurred naturally in different lineages over time and may be sufficient to explain much of the phenotypic variation in cortical field number and organization in different mammals.
Figures
Similar articles
-
Cross-modal reorganization of horizontal connectivity in auditory cortex without altering thalamocortical projections.J Neurosci. 1999 Sep 15;19(18):7940-50. doi: 10.1523/JNEUROSCI.19-18-07940.1999. J Neurosci. 1999. PMID: 10479695 Free PMC article.
-
Phenotypic diversity is the cornerstone of evolution: variation in cortical field size within short-tailed opossums.J Comp Neurol. 2006 Dec 20;499(6):990-9. doi: 10.1002/cne.21156. J Comp Neurol. 2006. PMID: 17072834
-
Thalamocortical projections in the reeler mutant mouse.J Comp Neurol. 1983 Sep 10;219(2):182-202. doi: 10.1002/cne.902190205. J Comp Neurol. 1983. PMID: 6194186
-
Development and plasticity of cortical areas and networks.Nat Rev Neurosci. 2001 Apr;2(4):251-62. doi: 10.1038/35067562. Nat Rev Neurosci. 2001. PMID: 11283748 Review.
-
Map transfer from the thalamus to the neocortex: inputs from the barrel field.Semin Cell Dev Biol. 2014 Nov;35:147-55. doi: 10.1016/j.semcdb.2014.07.005. Epub 2014 Jul 11. Semin Cell Dev Biol. 2014. PMID: 25020201 Review.
Cited by
-
A lifespan analysis of intraneocortical connections and gene expression in the mouse I.Cereb Cortex. 2011 Jun;21(6):1311-30. doi: 10.1093/cercor/bhq212. Epub 2010 Nov 8. Cereb Cortex. 2011. PMID: 21060110 Free PMC article.
-
Reorganization after pre- and perinatal brain lesions.J Anat. 2010 Oct;217(4):469-74. doi: 10.1111/j.1469-7580.2010.01262.x. J Anat. 2010. PMID: 20649910 Free PMC article. Review.
-
A mouse model of higher visual cortical function.Curr Opin Neurobiol. 2014 Feb;24(1):28-33. doi: 10.1016/j.conb.2013.08.009. Epub 2013 Sep 8. Curr Opin Neurobiol. 2014. PMID: 24492075 Free PMC article. Review.
-
Expansion and contraction of resource allocation in sensory bottlenecks.Elife. 2022 Aug 4;11:e70777. doi: 10.7554/eLife.70777. Elife. 2022. PMID: 35924884 Free PMC article.
-
Evolution of mammalian sensorimotor cortex: thalamic projections to parietal cortical areas in Monodelphis domestica.Front Neuroanat. 2015 Jan 7;8:163. doi: 10.3389/fnana.2014.00163. eCollection 2014. Front Neuroanat. 2015. PMID: 25620915 Free PMC article.
References
-
- Aitkin LM, Irvine DRF, Nelson JE, Merzenich MM, Clarey JC. Frequency representation in the auditory midbrain and forebrain of a marsupial, the northern native cat (Dasyurus hallucatus). Brain Behav Evol. 1986;29:17–28. - PubMed
-
- Brodmann K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. J. A. Barth; Leipzig: 1909.
-
- Bulfone A, Smiga SM, Shimamura K, Peterson A, Puelles L, Rubenstein JLR. T-Brain-1: a homolog of Brachyury whose expression defines molecularly distinct domains within the cerebral cortex. Neuron. 1995;15:63–78. - PubMed
-
- Carroll EW, Wong-Riley MTT. Quantitative light and electron microscopic analysis of cytochrome oxidase-rich zones in the striate cortex of the squirrel monkey. J Comp Neurol. 1984;222:1–17. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical