

Multiple components of the HSP90 chaperone complex function in regulation of heat shock factor 1 In vivo
- PMID: 10567529
- PMCID: PMC84888
- DOI: 10.1128/MCB.19.12.8033
Multiple components of the HSP90 chaperone complex function in regulation of heat shock factor 1 In vivo
Abstract
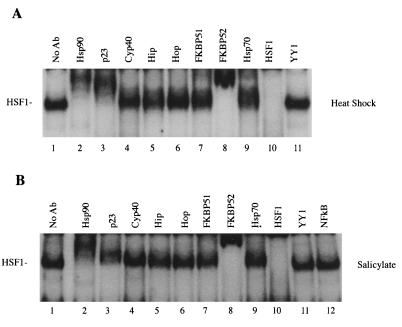
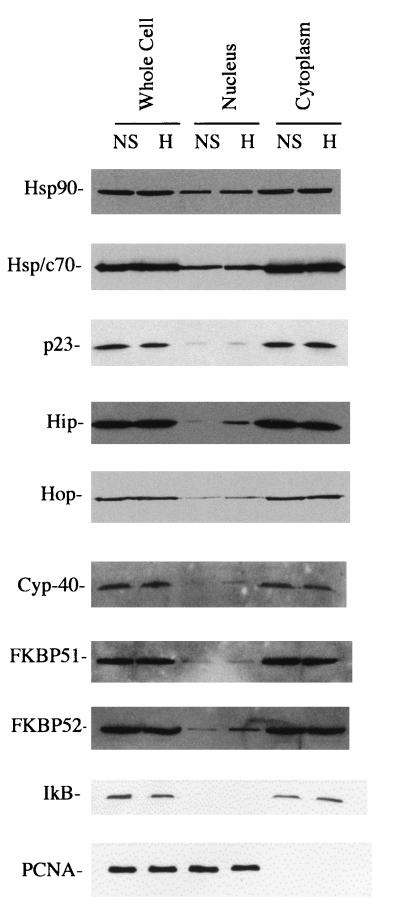
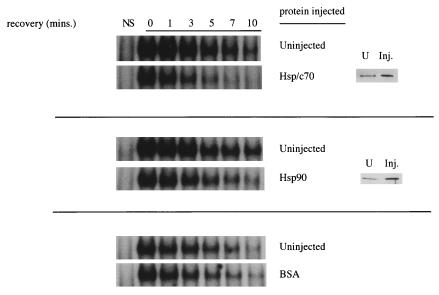
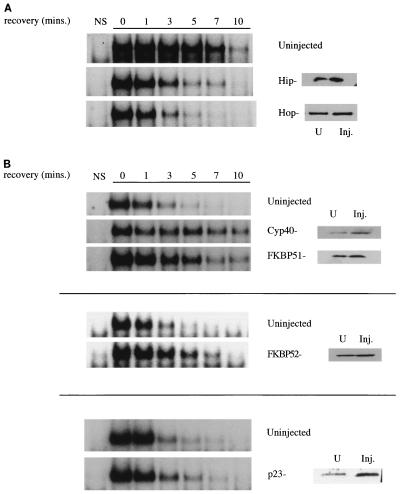
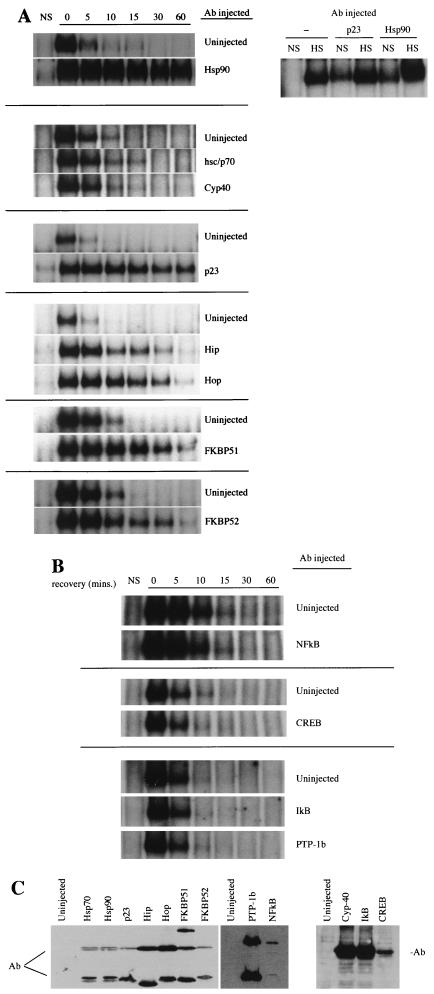
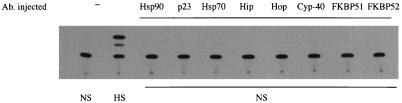
Rapid and transient activation of heat shock genes in response to stress is mediated in eukaryotes by the heat shock transcription factor HSF1. It is well established that cells maintain a dynamic equilibrium between inactive HSF1 monomers and transcriptionally active trimers, but little is known about the mechanism linking HSF1 to reception of various stress stimuli or the factors controlling oligomerization. Recent reports have revealed that HSP90 regulates key steps in the HSF1 activation-deactivation process. Here, we tested the hypothesis that components of the HSP90 chaperone machine, known to function in the folding and maturation of steroid receptors, might also participate in HSF1 regulation. Mobility supershift assays using antibodies against chaperone components demonstrate that active HSF1 trimers exist in a heterocomplex with HSP90, p23, and FKBP52. Functional in vivo experiments in Xenopus oocytes indicate that components of the HSF1 heterocomplex, as well as other components of the HSP90 cochaperone machine, are involved in regulating oligomeric transitions. Elevation of the cellular levels of cochaperones affected the time of HSF1 deactivation during recovery: attenuation was delayed by immunophilins, and accelerated by HSP90, Hsp/c70, Hip, or Hop. In immunotargeting experiments with microinjected antibodies, disruption of HSP90, Hip, Hop, p23, FKBP51, and FKBP52 delayed attenuation. In addition, HSF1 was activated under nonstress conditions after immunotargeting of HSP90 and p23, evidence that these proteins remain associated with HSF1 monomers and function in their repression in vivo. The remarkable similarity of HSF1 complex chaperones identified here (HSP90, p23, and FKBP52) and components in mature steroid receptor complexes suggests that HSF1 oligomerization is regulated by a foldosome-type mechanism similar to steroid receptor pathways. The current evidence leads us to propose a model in which HSF1, HSP90 and p23 comprise a core heterocomplex required for rapid conformational switching through interaction with a dynamic series of HSP90 subcomplexes.
Figures
References
-
- Abravaya K A, Myers M, Murphy S P, Morimoto R I. The human heat shock protein hsp70 interacts with HSF, the transcription factor that regulates heat shock gene expression. Genes Dev. 1992;6:1153–1164. - PubMed
-
- Ananthan J, Goldberg A L, Voellmy R. Abnormal proteins serve as eukaryotic stress signals and trigger the activation of heat shock genes. Science. 1986;232:522–525. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous