

Yap1p activates gene transcription in an oxidant-specific fashion
- PMID: 10567555
- PMCID: PMC84914
- DOI: 10.1128/MCB.19.12.8302
Yap1p activates gene transcription in an oxidant-specific fashion
Abstract
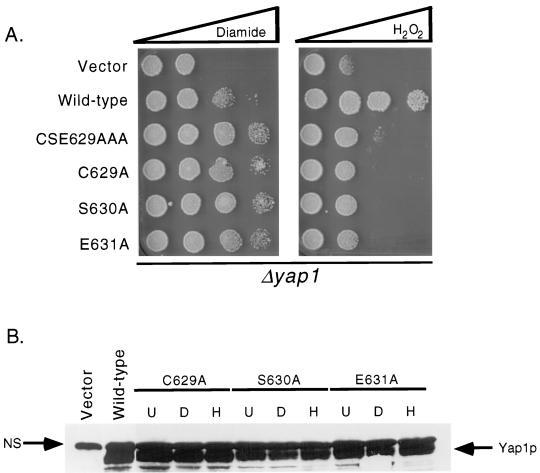
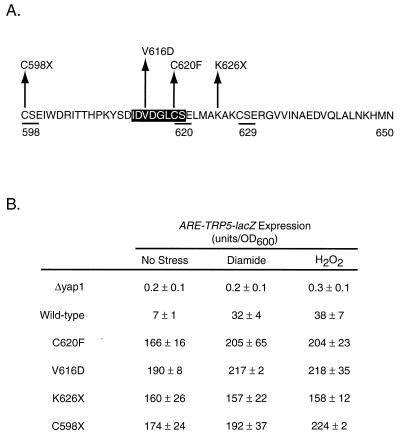
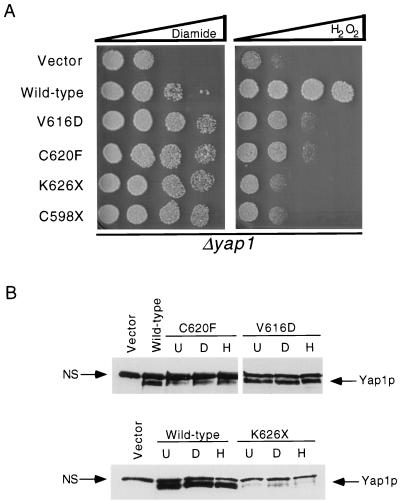
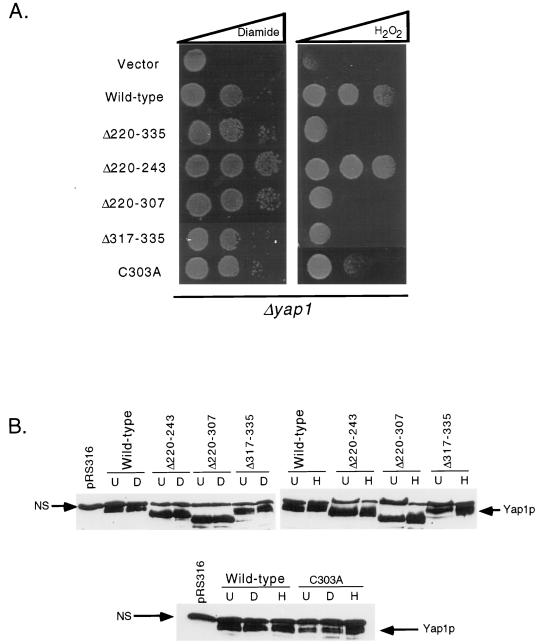
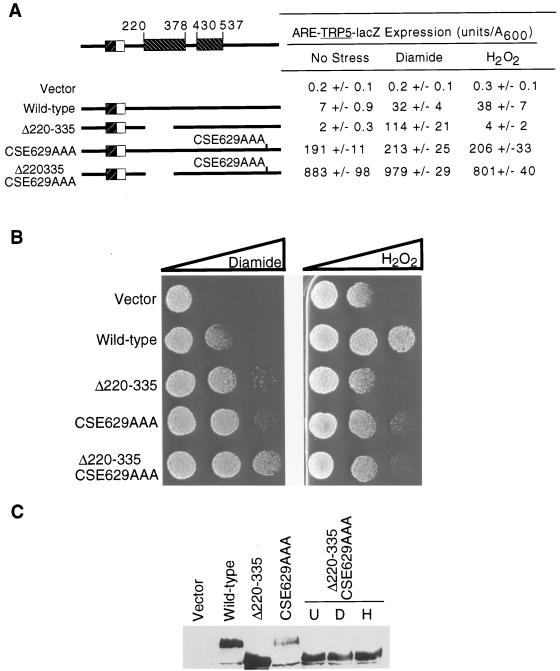
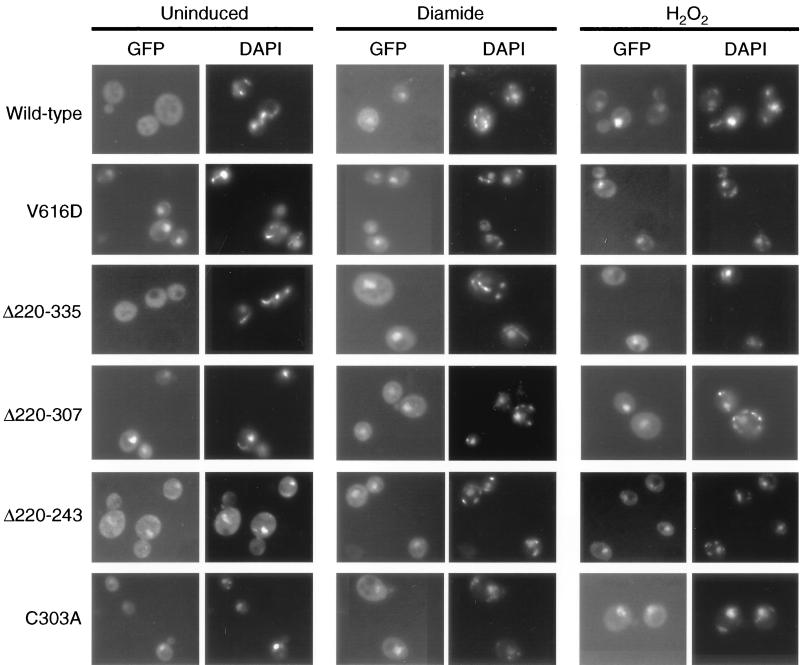
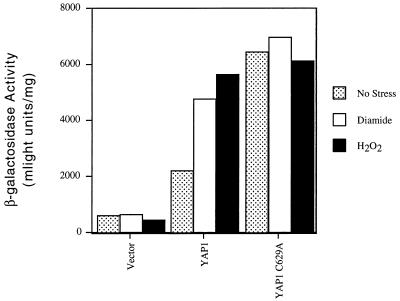
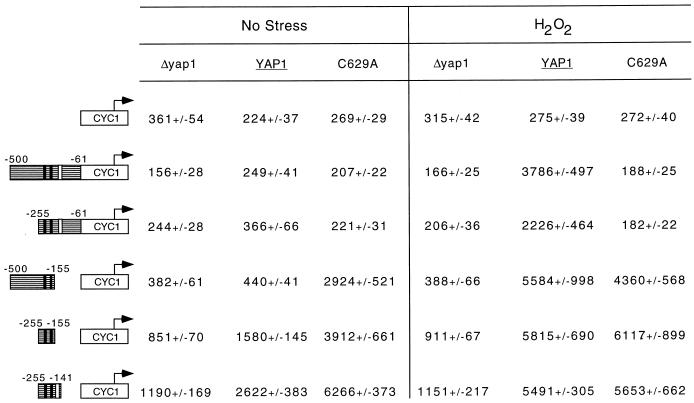
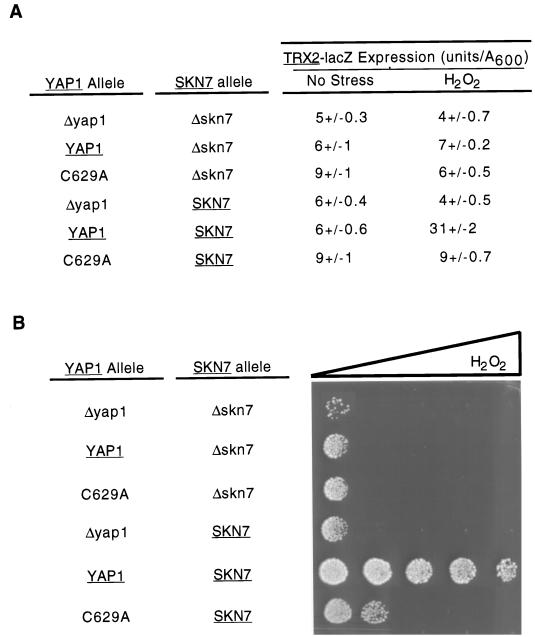
Positive regulation of gene expression by the yeast Saccharomyces cerevisiae transcription factor Yap1p is required for normal tolerance of oxidative stress elicited by the redox-active agents diamide and H(2)O(2). Several groups have provided evidence that a cluster of cysteine residues in the extreme C terminus of the factor are required for normal modulation of Yap1p by oxidant challenge. Deletion of this C-terminal cysteine-rich domain (c-CRD) produces a protein that is highly active under both stressed and nonstressed conditions and is constitutively located in the nucleus. We have found that a variety of different c-CRD mutant proteins are hyperactive in terms of their ability to confer diamide tolerance to cells but fail to provide even normal levels of H(2)O(2) resistance. Although the c-CRD mutant forms of Yap1p activate an artificial Yap1p-responsive gene to the same high level in the presence of either diamide or H(2)O(2), these mutant factors confer hyperresistance to diamide but hypersensitivity to H(2)O(2). To address this discrepancy, we have examined the ability of c-CRD mutant forms of Yap1p to activate expression of an authentic target gene required for H(2)O(2) tolerance, TRX2. When assayed in the presence of c-CRD mutant forms of Yap1p, a TRX2-lacZ fusion gene fails to induce in response to H(2)O(2). We have also identified a second cysteine-rich domain, in the N terminus (n-CRD), that is required for H(2)O(2) but not diamide resistance and influences the localization of the protein. These data are consistent with the idea that the function of Yap1p is different at promoters of loci involved in H(2)O(2) tolerance from promoters of genes involved in diamide resistance.
Figures


References
-
- Coleman S T, Tseng E, Moye-Rowley W S. Saccharomyces cerevisiae basic region-leucine zipper protein regulatory networks converge at the ATR1 structural gene. J Biol Chem. 1997;272:23224–23230. - PubMed
-
- Collinson L P, Dawes I W. Inducibility of the response of yeast cells to peroxide stress. J Gen Microbiol. 1992;138:329–335. - PubMed
-
- Grant C M, Collinson L P, Roe J-H, Dawes I W. Yeast glutathione reductase is required for protection against oxidative stress and is a target gene for yAP-1 transcriptional regulation. Mol Microbiol. 1996;21:739–746. - PubMed
-
- Grant C M, MacIver F H, Dawes I W. Glutathione is an essential metabolite required for resistance to oxidative stress in the yeast Saccharomyces cerevisiae. Curr Genet. 1996;29:511–515. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases