

Non-AUG initiation of AGAMOUS mRNA translation in Arabidopsis thaliana
- PMID: 10567575
- PMCID: PMC84964
- DOI: 10.1128/MCB.19.12.8505
Non-AUG initiation of AGAMOUS mRNA translation in Arabidopsis thaliana
Abstract
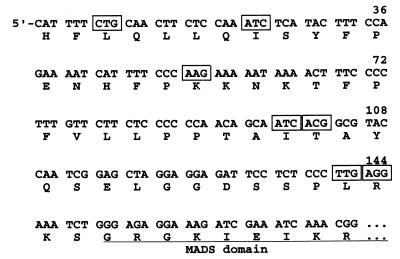
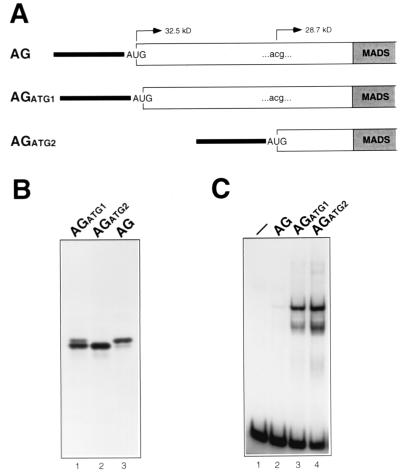
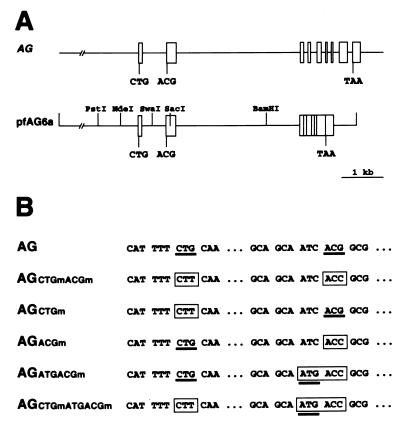
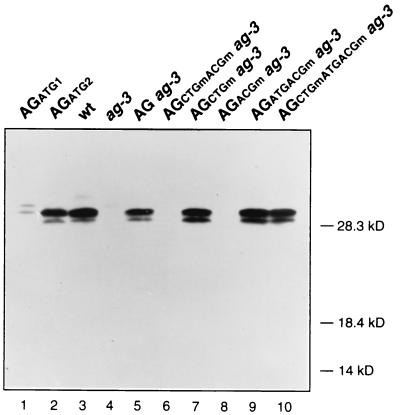
The MADS box organ identity gene AGAMOUS (AG) controls several steps during Arabidopsis thaliana flower development. AG cDNA contains an open reading frame that lacks an ATG triplet to function as the translation initiation codon, and the actual amino terminus of the AG protein remains uncharacterized. We have considered the possibility that AG translation can be initiated at a non-AUG codon. Two possible non-AUG initiation codons, CUG and ACG, are present in the 5' region of AG mRNA preceding the highly conserved MADS box sequence. We prepared a series of AG genomic constructs in which these codons are mutated and assayed their activity in phenotypic rescue experiments by introducing them as transgenes into ag mutant plants. Alteration of the CTG codon to render it unsuitable for acting as a translation initiation site does not affect complementation of the ag-3 mutation in transgenic plants. However, a similar mutation of the downstream ACG codon prevents the rescue of the ag-3 mutant phenotype. Conversely, if an ATG is introduced immediately 5' to the disrupted ACG codon, the resulting construct fully complements the ag-3 mutation. The AG protein synthesized in vitro by initiating translation at the ACG position is active in DNA binding and is of the same size as the AG protein detected from floral tissues, whereas AG polypeptides with additional amino-terminal residues do not appear to bind DNA. These results indicate that translation of AG is initiated exclusively at an ACG codon and prove that non-AUG triplets may be efficiently used as the sole translation initiation site in some plant cellular mRNAs.
Figures


References
-
- Acland P, Dixon M, Peters G, Dickson C. Subcellular fate of the Int-2 oncoprotein is determined by choice of initiation codon. Nature. 1990;343:662–665. - PubMed
-
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci. 1993;316:1194–1199.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases