

Functional analysis of the DXPas34 locus, a 3' regulator of Xist expression
- PMID: 10567576
- PMCID: PMC84967
- DOI: 10.1128/MCB.19.12.8513
Functional analysis of the DXPas34 locus, a 3' regulator of Xist expression
Abstract
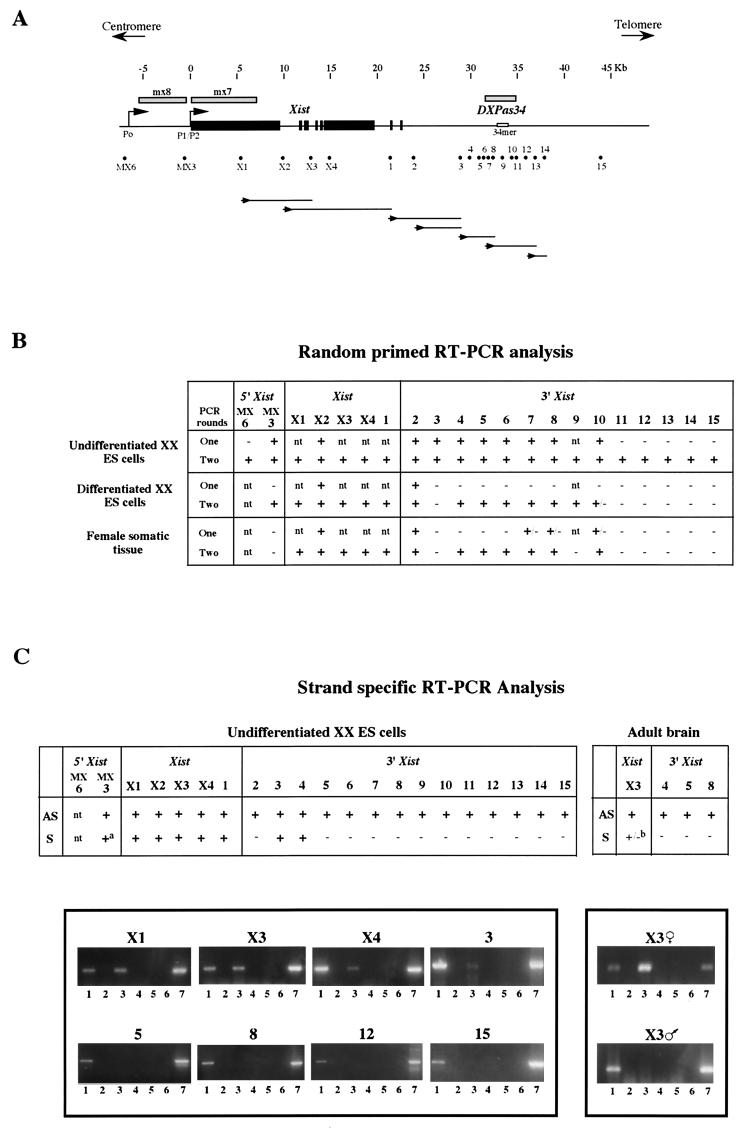
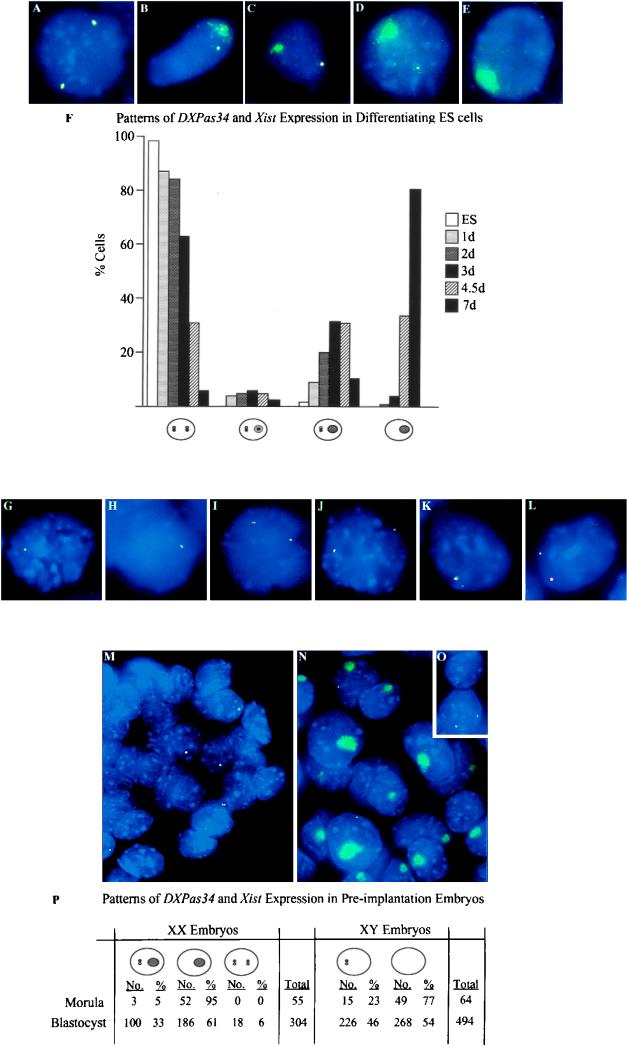
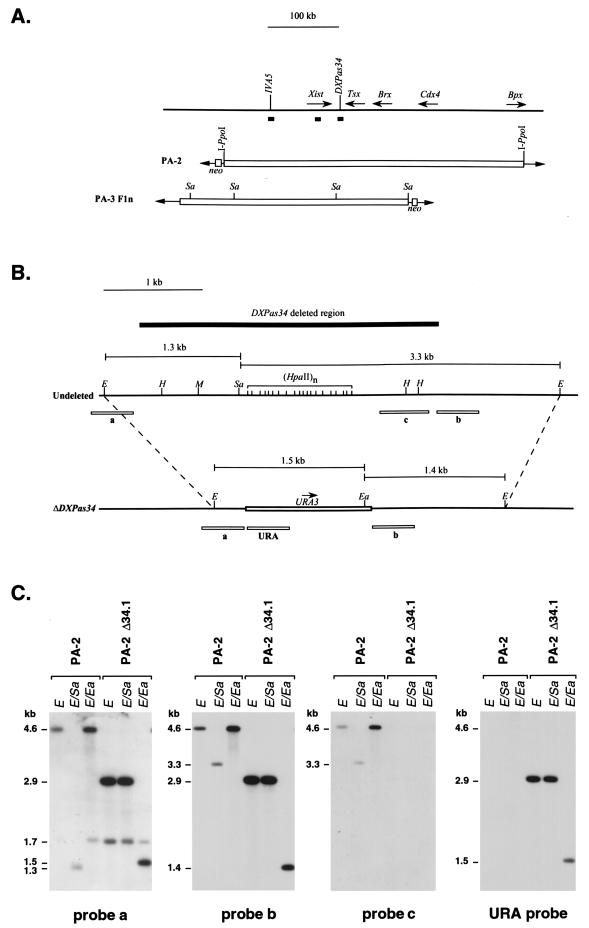
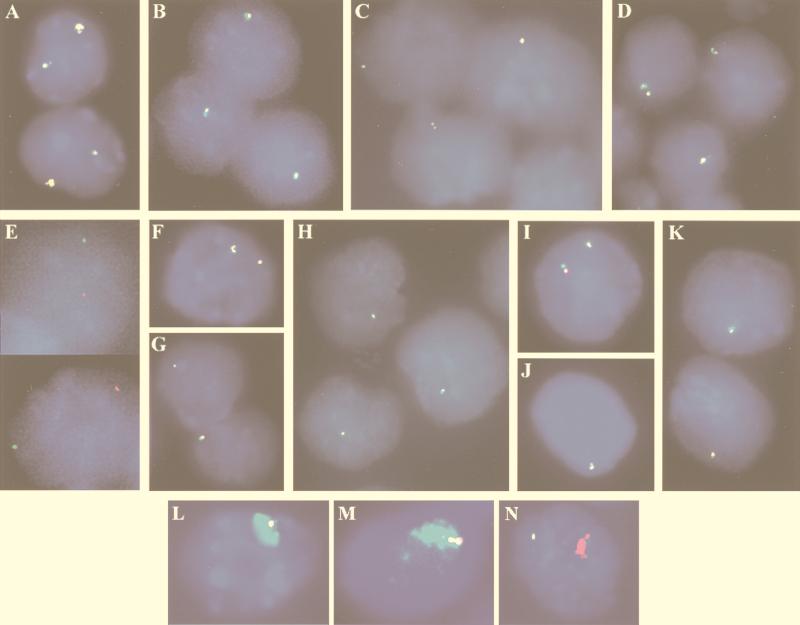
X inactivation in female mammals is controlled by a key locus on the X chromosome, the X-inactivation center (Xic). The Xic controls the initiation and propagation of inactivation in cis. It also ensures that the correct number of X chromosomes undergo inactivation (counting) and determines which X chromosome becomes inactivated (choice). The Xist gene maps to the Xic region and is essential for the initiation of X inactivation in cis. Regulatory elements of X inactivation have been proposed to lie 3' to Xist. One such element, lying 15 kb downstream of Xist, is the DXPas34 locus, which was first identified as a result of its hypermethylation on the active X chromosome and the correlation of its methylation level with allelism at the X-controlling element (Xce), a locus known to affect choice. In this study, we have tested the potential function of the DXPas34 locus in Xist regulation and X-inactivation initiation by deleting it in the context of large Xist-containing yeast artificial chromosome transgenes. Deletion of DXPas34 eliminates both Xist expression and antisense transcription present in this region in undifferentiated ES cells. It also leads to nonrandom inactivation of the deleted transgene upon differentiation. DXPas34 thus appears to be a critical regulator of Xist activity and X inactivation. The expression pattern of DXPas34 during early embryonic development, which we report here, further suggests that it could be implicated in the regulation of imprinted Xist expression.
Figures
References
-
- Avner P. X chromosome inactivation: Xce and the candidate region for the X inactivation centre. In: Russo E, Riggs A, Martienssen R, editors. Epigenetic mechanisms of gene regulation. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1996. pp. 249–265.
-
- Avner P, Prissette M, Arnaud D, Courtier B, Cecchi C, Heard E. Molecular correlates of the murine Xce locus. Genet Res. 1998;72:217–224. - PubMed
-
- Beddington R S P. Isolation, culture and manipulation of postimplantation mouse embryos. Oxford, England: IRL Press; 1987.
-
- Borsani B, Tonlorenzi R, Simmler M-C, Dandolo L, Arnaud D, Capra V, Grompe M, Pizzuti A, Muzni D, Lawrence C, Willard H F, Avner P, Ballabio A. Characterization of a murine gene expressed from the inactive X chromosome. Nature. 1991;351:325–329. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases