

Conservation of a gliding motility and cell invasion machinery in Apicomplexan parasites
- PMID: 10579715
- PMCID: PMC2169348
- DOI: 10.1083/jcb.147.5.937
Conservation of a gliding motility and cell invasion machinery in Apicomplexan parasites
Abstract
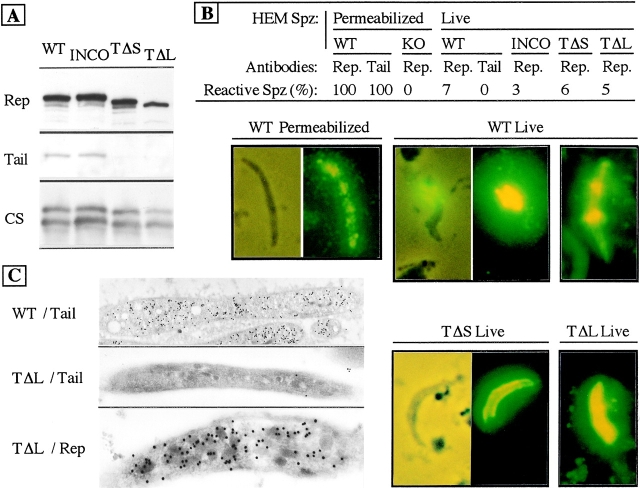
Most Apicomplexan parasites, including the human pathogens Plasmodium, Toxoplasma, and Cryptosporidium, actively invade host cells and display gliding motility, both actions powered by parasite microfilaments. In Plasmodium sporozoites, thrombospondin-related anonymous protein (TRAP), a member of a group of Apicomplexan transmembrane proteins that have common adhesion domains, is necessary for gliding motility and infection of the vertebrate host. Here, we provide genetic evidence that TRAP is directly involved in a capping process that drives both sporozoite gliding and cell invasion. We also demonstrate that TRAP-related proteins in other Apicomplexa fulfill the same function and that their cytoplasmic tails interact with homologous partners in the respective parasite. Therefore, a mechanism of surface redistribution of TRAP-related proteins driving gliding locomotion and cell invasion is conserved among Apicomplexan parasites.
Figures


Similar articles
-
TREP, a novel protein necessary for gliding motility of the malaria sporozoite.Int J Parasitol. 2009 Mar;39(4):489-96. doi: 10.1016/j.ijpara.2008.10.004. Epub 2008 Oct 25. Int J Parasitol. 2009. PMID: 19000911
-
A conserved molecular motor drives cell invasion and gliding motility across malaria life cycle stages and other apicomplexan parasites.J Biol Chem. 2006 Feb 24;281(8):5197-208. doi: 10.1074/jbc.M509807200. Epub 2005 Dec 1. J Biol Chem. 2006. PMID: 16321976
-
Microneme proteins: structural and functional requirements to promote adhesion and invasion by the apicomplexan parasite Toxoplasma gondii.Int J Parasitol. 2001 Oct;31(12):1293-302. doi: 10.1016/s0020-7519(01)00257-0. Int J Parasitol. 2001. PMID: 11566297 Review.
-
TRAP is necessary for gliding motility and infectivity of plasmodium sporozoites.Cell. 1997 Aug 8;90(3):511-22. doi: 10.1016/s0092-8674(00)80511-5. Cell. 1997. PMID: 9267031
-
[The molecular mechanisms of erythrocyte invasion of Plasmodium spp. as a model organism of apicomplexan protozoa].Turkiye Parazitol Derg. 2010;34(4):203-6. doi: 10.5152/tpd.2010.15. Turkiye Parazitol Derg. 2010. PMID: 21391195 Review. Turkish.
Cited by
-
Regulation and use of the extracellular matrix by Trypanosoma cruzi during early infection.Front Immunol. 2012 Nov 6;3:337. doi: 10.3389/fimmu.2012.00337. eCollection 2012. Front Immunol. 2012. PMID: 23133440 Free PMC article.
-
Structural and functional dissection of the adhesive domains of Plasmodium falciparum thrombospondin-related anonymous protein (TRAP).Biochem J. 2004 May 1;379(Pt 3):815-22. doi: 10.1042/BJ20031500. Biochem J. 2004. PMID: 14741048 Free PMC article.
-
The claudin-like apicomplexan microneme protein is required for gliding motility and infectivity of Plasmodium sporozoites.PLoS Pathog. 2023 Mar 16;19(3):e1011261. doi: 10.1371/journal.ppat.1011261. eCollection 2023 Mar. PLoS Pathog. 2023. PMID: 36928686 Free PMC article.
-
Identification and Application of Epitopes in EtMIC1 of Eimeria tenella Recognized by the Monoclonal Antibodies 1-A1 and 1-H2.Infect Immun. 2019 Oct 18;87(11):e00596-19. doi: 10.1128/IAI.00596-19. Print 2019 Nov. Infect Immun. 2019. PMID: 31427452 Free PMC article.
-
Cytoskeleton of apicomplexan parasites.Microbiol Mol Biol Rev. 2002 Mar;66(1):21-38; table of contents. doi: 10.1128/MMBR.66.1.21-38.2002. Microbiol Mol Biol Rev. 2002. PMID: 11875126 Free PMC article. Review.
References
-
- Aikawa M., Atkinson C.T. Immunoelectron microscopy of parasites. Adv. Parasitol. 1990;29:151–214. - PubMed
-
- Carruthers V.B., Sibley L.D. Sequential protein secretion from three distinct organelles of Toxoplasma gondii accompanies invasion of human fibroblasts. Eur. J. Cell Biol. 1997;73:114–123. - PubMed
-
- Dobrowolski J.M., Sibley L.D. Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell. 1996;84:933–939. - PubMed
-
- Dobrowolski J.M., Carruthers V.B., Sibley L.D. Participation of myosin in gliding motility and host cell invasion by Toxoplasma gondii . Mol. Microbiol. 1997;26:163–173. - PubMed
