

Peroxynitrite is a positive inotropic agent in atrial and ventricular fibres of the frog heart
- PMID: 10581309
- PMCID: PMC2269669
- DOI: 10.1111/j.1469-7793.1999.00375.x
Peroxynitrite is a positive inotropic agent in atrial and ventricular fibres of the frog heart
Abstract
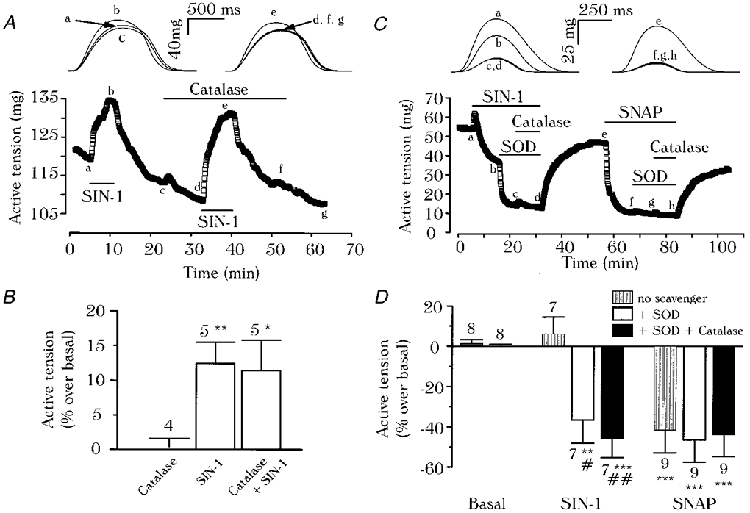
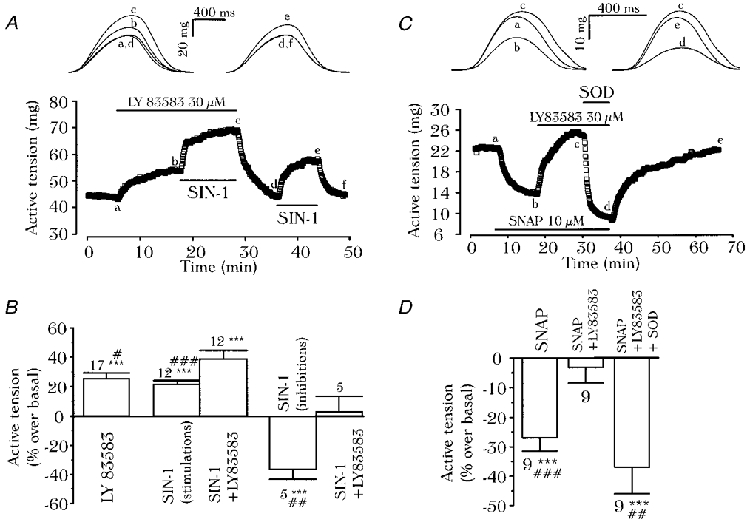
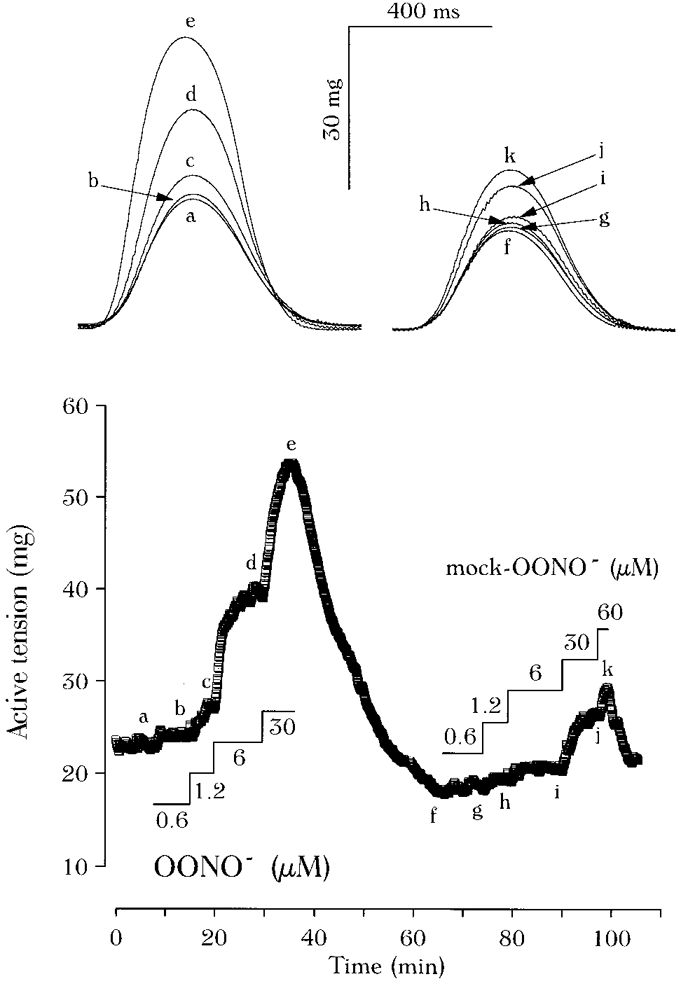
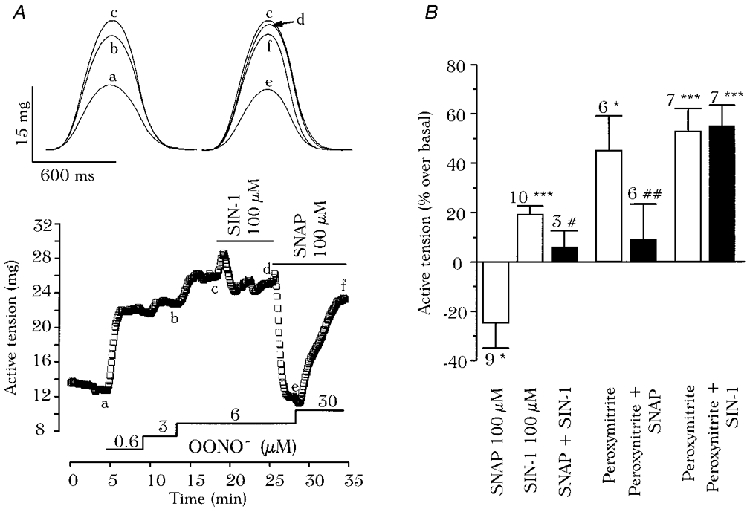
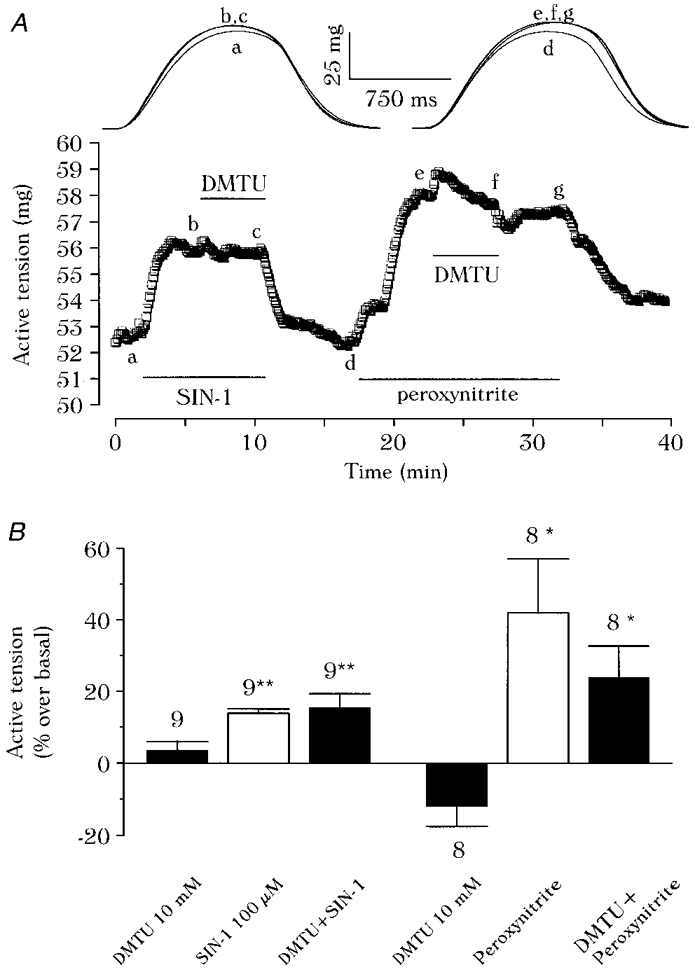
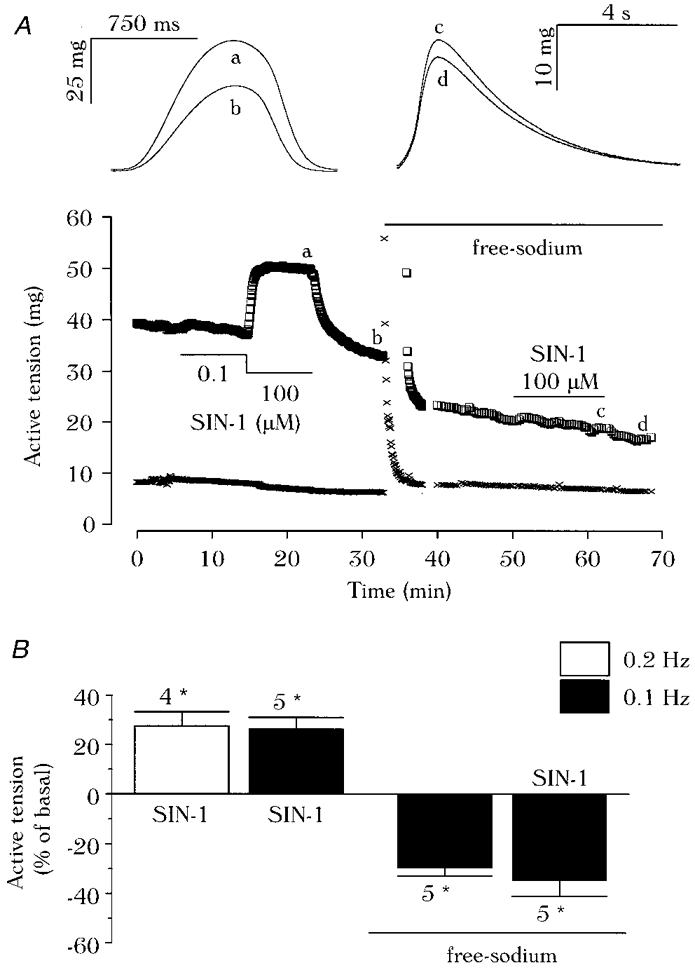
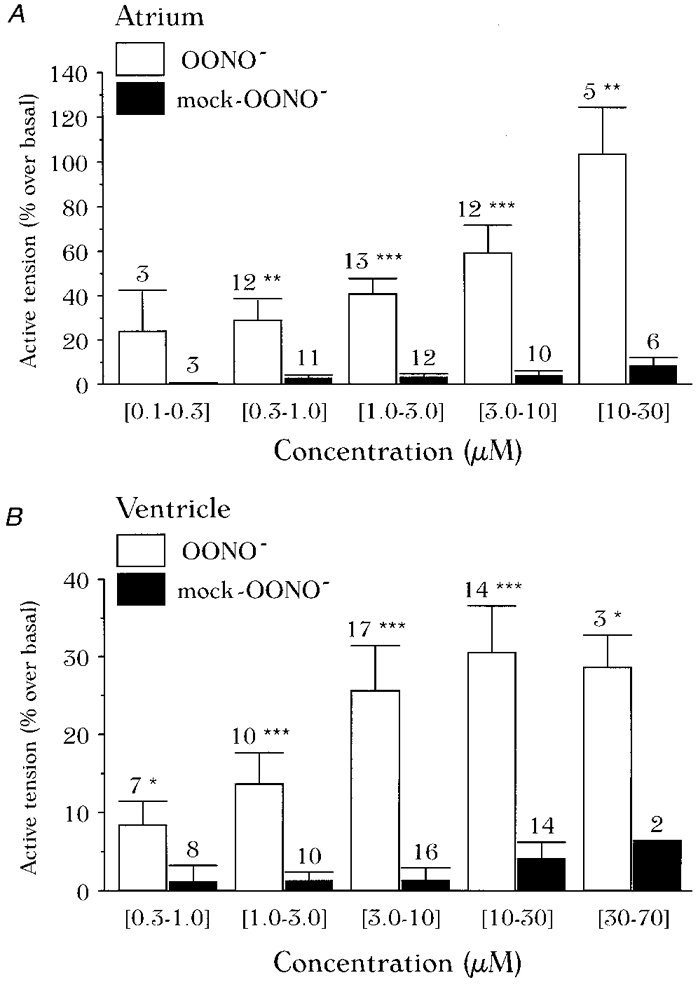
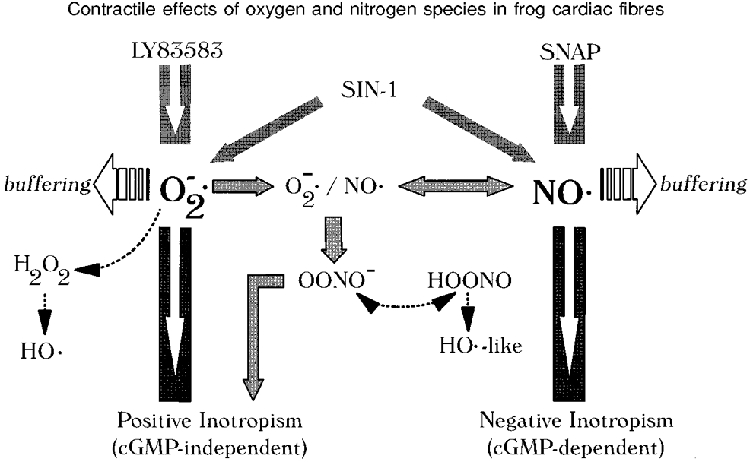
1. We report opposite inotropic effects of NO donors in frog cardiac fibres. The negative effect, elicited by either 3-morpholino-sydnonimine (SIN-1) or S-nitroso-N-acetyl-penicillamine (SNAP), involved cyclic GMP (cGMP) production. However, SIN-1, unlike SNAP, could elicit a positive effect, in a superoxide dismutase (SOD)-sensitive manner. SIN-1, unlike SNAP, can release both NO and superoxide anion, the precursors of peroxynitrite (OONO-). The role of these messengers was examined. 2. Catalase did not reduce the positive inotropic effect of SIN-1. Thus, a conversion of superoxide anion into hydrogen peroxide was not involved in this effect. In addition, catalase did not modify the negative effects of SIN-1 plus SOD, or SNAP plus SOD. 3. LY 83583, a superoxide anion generator, elicited a positive inotropic effect, like SIN-1. The effect of LY 83583 was additive to the negative effects of SIN-1 or SNAP, and to the positive effect of SIN-1. Thus, superoxide anion generation, per se, did not account for the positive effect of SIN-1. 4. Authentic peroxynitrite (OONO-), but not mock-OONO- (negative control plus decomposed OONO-), exerted a dramatic positive inotropic effect in cardiac fibres. The effect of OONO- was larger in atrial fibres, as compared with ventricular fibres. 5. The positive effect of OONO- was not additive with that of SIN-1, suggesting a common mechanism of action. In contrast, the effects of either OONO- or SIN-1 were additive with the negative inotropic effect of SNAP. Furthermore, the effect of OONO-, like that of SIN-1, was not antagonized by 1H-[1,2,4]xidiazolo[4, 3-a]quinoxaline-1-one (ODQ; 10 microM), the guanylyl cyclase inhibitor. 6. The positive inotropic effects of SIN-1 and OONO- were not modified by hydroxyl radical scavengers, such as dimethyl-thio-urea (DMTU; 10 mM). 7. The positive inotropic effect of SIN-1 (100 microM) was abolished in sodium-free solutions, a treatment that eliminates the activity of the sodium-calcium exchanger. In contrast, the effect of SIN-1 was unchanged by a potassium channel inhibitor (tetraethyl-ammonium, 20 mM), or a sodium-potassium pump inhibitor (ouabain 10 microM). 8. We conclude that OONO- is a positive inotropic agent in frog cardiac fibres. The generation of OONO- accounts for the positive inotropic effect of SIN-1. OONO- itself was responsible for the positive inotropic effect, and appeared to modulate the activity of the sodium-calcium exchanger.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources