

NOS2-derived nitric oxide regulates the size, quantity and quality of granuloma formation in Mycobacterium avium-infected mice without affecting bacterial loads
- PMID: 10583588
- PMCID: PMC2326941
- DOI: 10.1046/j.1365-2567.1999.00875.x
NOS2-derived nitric oxide regulates the size, quantity and quality of granuloma formation in Mycobacterium avium-infected mice without affecting bacterial loads
Abstract
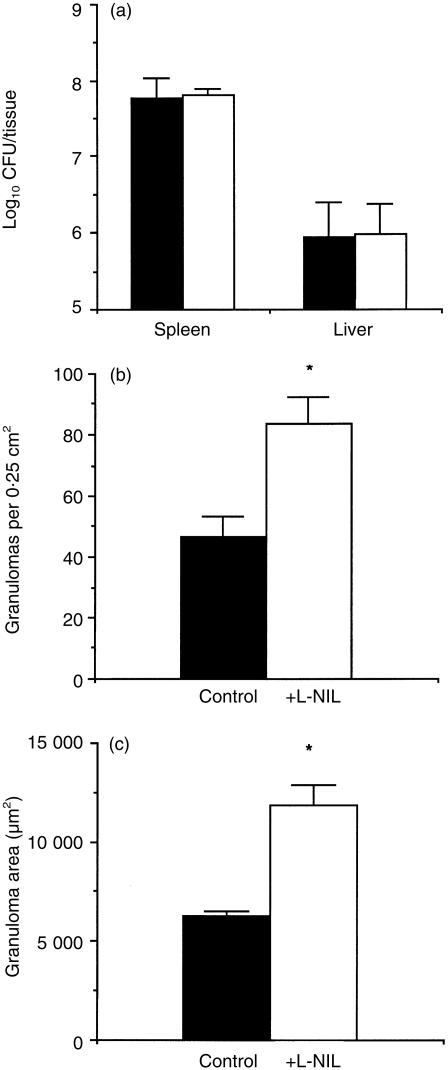
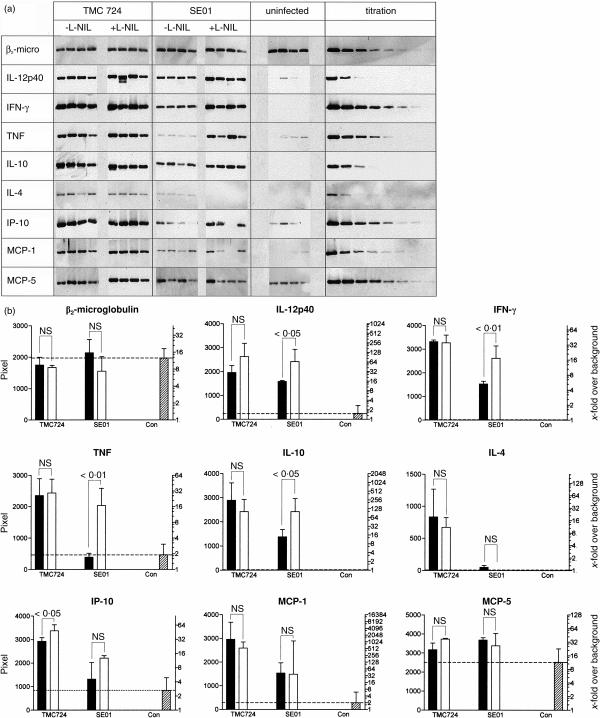
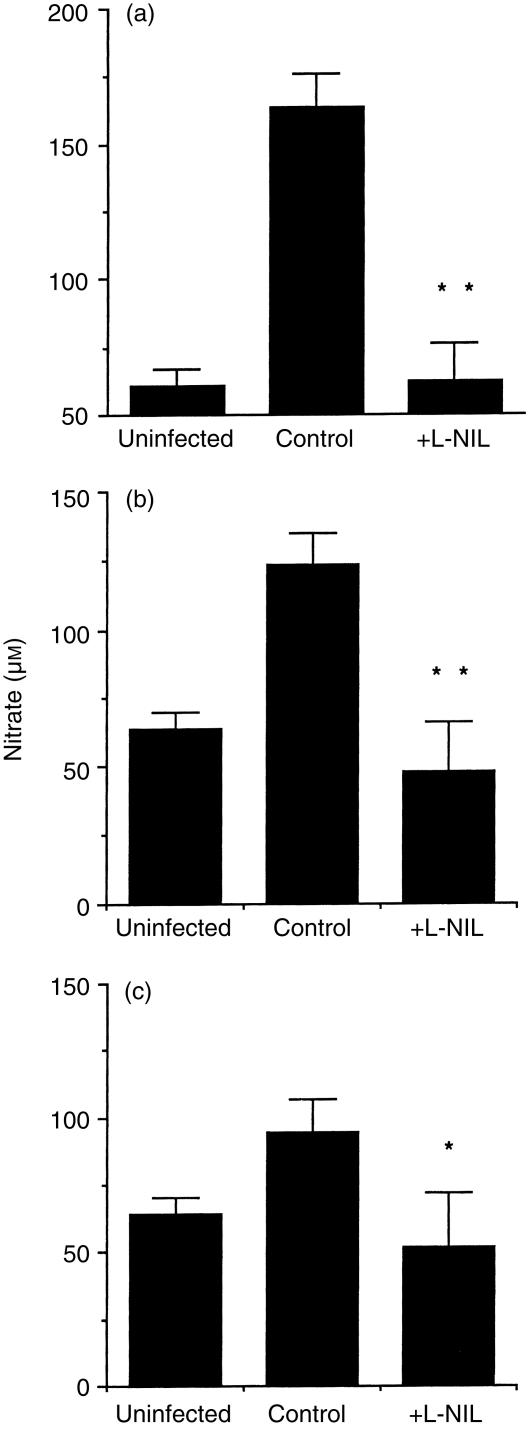
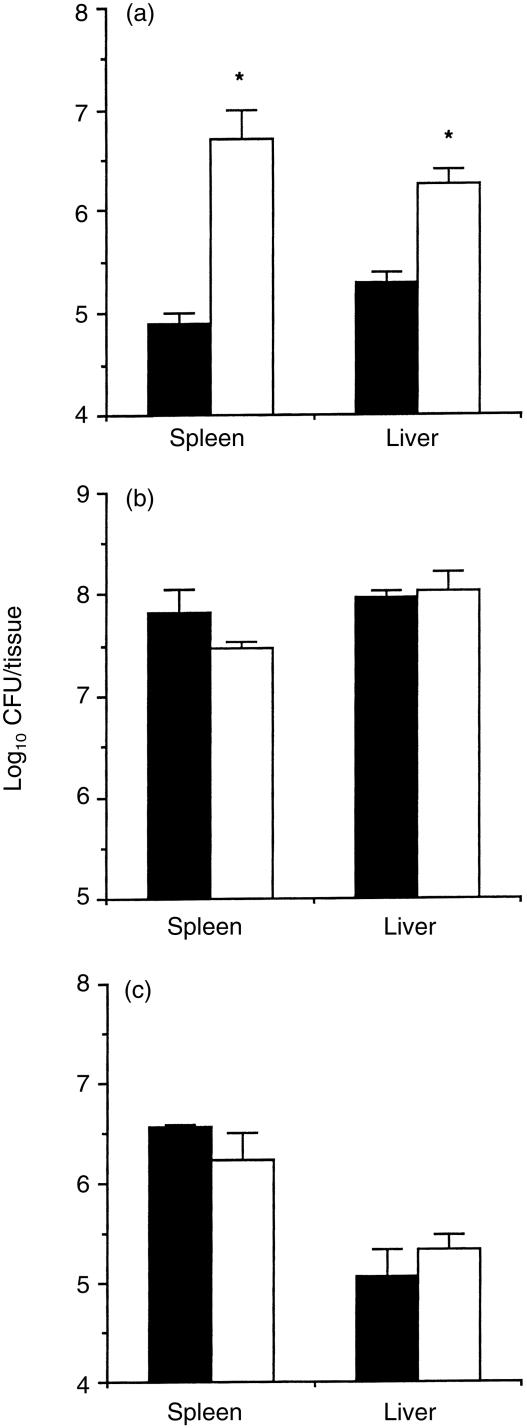
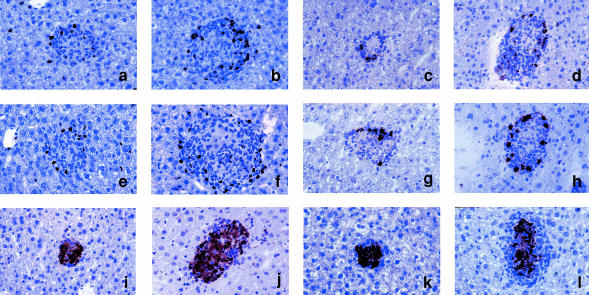
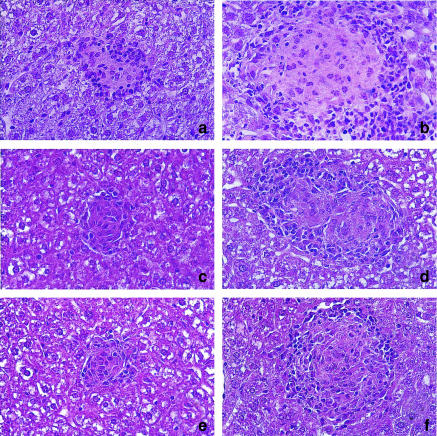
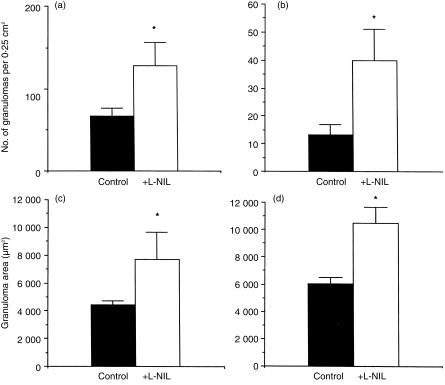
Granuloma formation in response to mycobacterial infections is associated with increased expression of inducible nitric oxide synthase (NOS2) within granuloma macrophages and increased levels of nitrate/nitrite in the sera of infected mice. Continuous treatment with 5 mm or 10 mm l-N6-(1-imino-ethyl)-lysine (L-NIL), a selective NOS2-inhibitor, in acidified drinking water for up to 7 weeks consistently reduced infection-induced nitrate/nitrite to background levels in mycobacteria-infected BALB/c mice. Oral treatment with 5 mm L-NIL initiated at the time of infection significantly exacerbated growth of Mycobacterium tuberculosis, but had no effect on Mycobacterium avium colony-forming unit development in the liver, spleen and lungs of intravenously infected mice. In order to examine the role of nitric oxide in mycobacteria-induced granulomatous inflammation in the absence of any effect on the bacterial load, M. avium-infected mice were treated with 5 mm L-NIL from day 1 through 38 and the development of granulomatous lesions in the liver was assessed by histology, immunohistology and reverse-transcription-polymerase chain reaction (RT-PCR). Computer- and video-assisted morphometry performed at 4 and 7 weeks post-infection showed that treatment with L-NIL led to markedly increased number, cellularity and size of granulomatous lesions in infected mice regardless of the virulence of the M. avium isolate used for infection. Immunohistology of the liver revealed that in mice treated with L-NIL, the numbers of CD3+ T cells, CD21/35+ B cells, CD11b+ macrophages and RB6-8C5+ granulocytes associated with granulomatous lesions was increased. RT-PCR of the liver showed that in L-NIL-treated mice infected with M. avium, mRNA levels of tumour necrosis factor, interleukin-12p40, interferon-gamma, interleukin-10 and interferon-gamma-inducible protein-10 (IP-10) were up-regulated, while mRNA levels of interleukin-4, monocyte chemotactic protein-1 (MCP-1) and MCP-5 were similar to those in untreated control infected mice. When M. avium-infected mice were treated with 5 mm L-NIL between the 5th and 12th weeks of infection, similar changes in granuloma number and size were found in the absence of any effect on the bacterial load. These findings demonstrate that nitric oxide regulates the number, size and cellular composition of M. avium-induced granulomas independently of antibacterial effects by modulating the cytokine profile within infected tissues.
Figures
Similar articles
-
Fatal granuloma necrosis without exacerbated mycobacterial growth in tumor necrosis factor receptor p55 gene-deficient mice intravenously infected with Mycobacterium avium.Infect Immun. 1999 Jul;67(7):3571-9. doi: 10.1128/IAI.67.7.3571-3579.1999. Infect Immun. 1999. PMID: 10377141 Free PMC article.
-
Control of Mycobacterium fortuitum and Mycobacterium intracellulare infections with respect to distinct granuloma formations in livers of BALB/c mice.Mem Inst Oswaldo Cruz. 2010 Aug;105(5):642-8. doi: 10.1590/s0074-02762010000500007. Mem Inst Oswaldo Cruz. 2010. PMID: 20835610
-
Mycobacterium avium infection in CD14-deficient mice fails to substantiate a significant role for CD14 in antimycobacterial protection or granulomatous inflammation.Immunology. 2001 May;103(1):113-21. doi: 10.1046/j.1365-2567.2001.01214.x. Immunology. 2001. PMID: 11380699 Free PMC article.
-
Three-dimensional in vitro models of granuloma to study bacteria-host interactions, drug-susceptibility, and resuscitation of dormant mycobacteria.Biomed Res Int. 2014;2014:623856. doi: 10.1155/2014/623856. Epub 2014 May 21. Biomed Res Int. 2014. PMID: 24967387 Free PMC article. Review.
-
Heterogeneity in the granulomatous response to mycobacterial infection in patients with defined genetic mutations in the interleukin 12-dependent interferon-gamma production pathway.Int J Exp Pathol. 2002 Feb;83(1):1-20. doi: 10.1046/j.1365-2613.2002.00216.x. Int J Exp Pathol. 2002. PMID: 12059906 Free PMC article. Review.
Cited by
-
Regulation of granuloma fibrosis by nitric oxide during Mycobacterium avium experimental infection.Int J Exp Pathol. 2006 Aug;87(4):307-15. doi: 10.1111/j.1365-2613.2006.00487.x. Int J Exp Pathol. 2006. PMID: 16875496 Free PMC article.
-
Gamma interferon is essential for clearing Mycobacterium genavense infection.Infect Immun. 2000 Jun;68(6):3720-3. doi: 10.1128/IAI.68.6.3720-3723.2000. Infect Immun. 2000. PMID: 10816534 Free PMC article.
-
Characterization of membrane vesicles released by Mycobacterium avium in response to environment mimicking the macrophage phagosome.Future Microbiol. 2019 Mar;14(4):293-313. doi: 10.2217/fmb-2018-0249. Epub 2019 Feb 13. Future Microbiol. 2019. PMID: 30757918 Free PMC article.
-
Pathogenesis of Mycobacterium avium infection: typical responses to an atypical mycobacterium?Immunol Res. 2006;35(3):179-90. doi: 10.1385/IR:35:3:179. Immunol Res. 2006. PMID: 17172645 Review.
-
Persimmon-derived tannin has bacteriostatic and anti-inflammatory activity in a murine model of Mycobacterium avium complex (MAC) disease.PLoS One. 2017 Aug 21;12(8):e0183489. doi: 10.1371/journal.pone.0183489. eCollection 2017. PLoS One. 2017. PMID: 28827842 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
