

Adenovirus type 37 uses sialic acid as a cellular receptor
- PMID: 10590089
- PMCID: PMC111511
Adenovirus type 37 uses sialic acid as a cellular receptor
Abstract
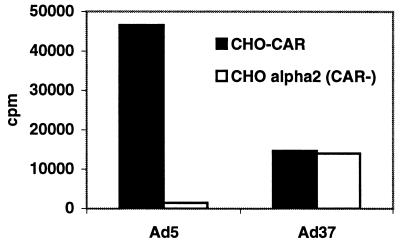
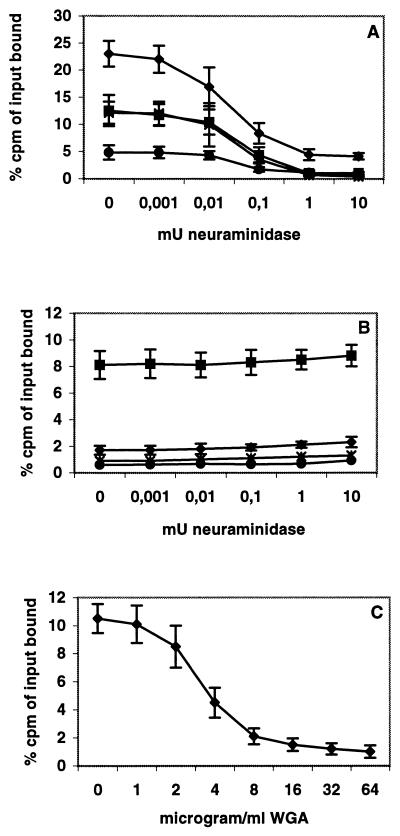
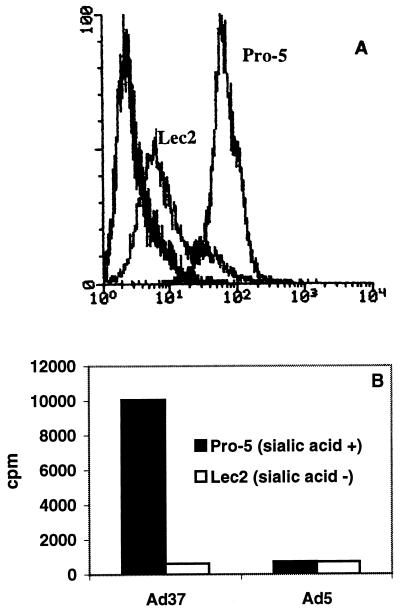
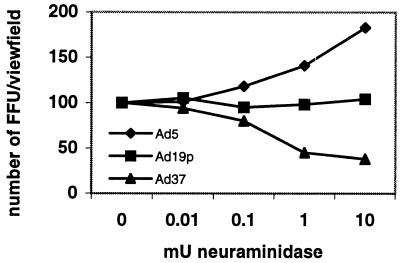
Two cellular receptors for adenovirus, coxsackievirus-adenovirus receptor (CAR) and major histocompatibility complex class I (MHC-I) alpha2, have recently been identified. In the absence of CAR, MHC-I alpha2 has been suggested to serve as a cellular attachment protein for subgenus C adenoviruses, while members from all subgenera except subgenus B have been shown to interact with CAR. We have found that adenovirus type 37 (Ad37) attachment to CAR-expressing CHO cells was no better than that to CHO cells lacking CAR expression, suggesting that CAR is not used by Ad37 during attachment. Instead, we have identified sialic acid as a third adenovirus receptor moiety. First, Ad37 attachment to both CAR-expressing CHO cells and MHC-I alpha2-expressing Daudi cells was sensitive to neuraminidase treatment, which eliminates sialic acid on the cell surface. Second, Ad37 attachment to sialic acid-expressing Pro-5 cells was more than 10-fold stronger than that to the Pro-5 subline Lec2, which is deficient in sialic acid expression. Third, neuraminidase treatment of A549 cells caused a 60% decrease in Ad37 replication in a fluorescent-focus assay. Moreover, the receptor sialoconjugate is most probably a glycoprotein rather than a ganglioside, since Ad37 attachment to sialic acid-expressing Pro-5 cells was sensitive to protease treatment. Ad37 attachment to Pro-5 cells occurs via alpha(2-->3)-linked sialic acid saccharides rather than alpha(2-->6)-linked ones, since (i) alpha(2-->3)-specific but not alpha(2-->6)-specific lectins blocked Ad37 attachment to Pro-5 cells and (ii) pretreatment of Pro-5 cells with alpha(2-->3)-specific neuraminidase resulted in decreased Ad37 binding. Taken together, these results suggest that, unlike Ad5, Ad37 makes use of alpha(2-->3)-linked sialic acid saccharides on glycoproteins for entry instead of using CAR or MHC-I alpha2.
Figures
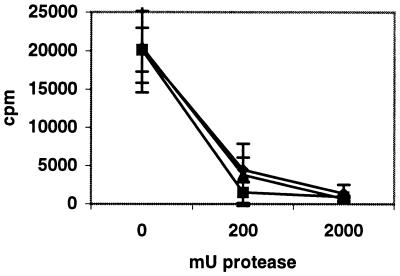
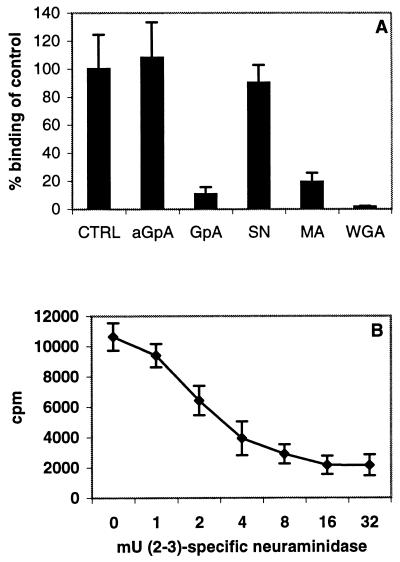
References
-
- Akusjärvi G, Aleström P, Pettersson M, Lager M, Jörnvall H, Pettersson U. The gene for adenovirus 2 hexon protein. J Biol Chem. 1984;25:13976–13979. - PubMed
-
- Araki-Sasaki K, Ohasi Y, Sasabe T, Hayashi K, Watanabe H, Tano Y, Handa H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig Ophthalmol Vis Sci. 1995;36:614–621. - PubMed
-
- Arcasoy S M, Latoche J, Gondor M, Watkins S C, Henderson R A, Hughey R, Finn O J, Pilewski J M. MUC1 and other sialoglycoconjugates inhibit adenovirus-mediated gene transfer to epithelial cells. Am J Respir Cell Mol Biol. 1997;17:422–435. - PubMed
-
- Arnberg N, Mei Y, Wadell G. Fiber genes of adenoviruses with tropism for the eye and the genital tract. Virology. 1997;227:239–244. - PubMed
-
- Basak S, Turner H, Parr S. Identification of a 40–42 kDa attachment polypeptide for canine parvovirus in A72 cells. Virology. 1994;205:7–16. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials