

Initial events in infectious salmon anemia virus infection: evidence for the requirement of a low-pH step
- PMID: 10590109
- PMCID: PMC111531
- DOI: 10.1128/jvi.74.1.218-227.2000
Initial events in infectious salmon anemia virus infection: evidence for the requirement of a low-pH step
Abstract
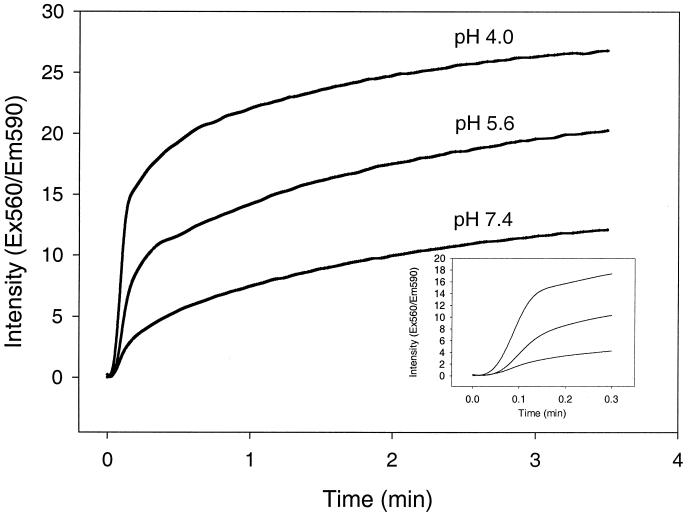
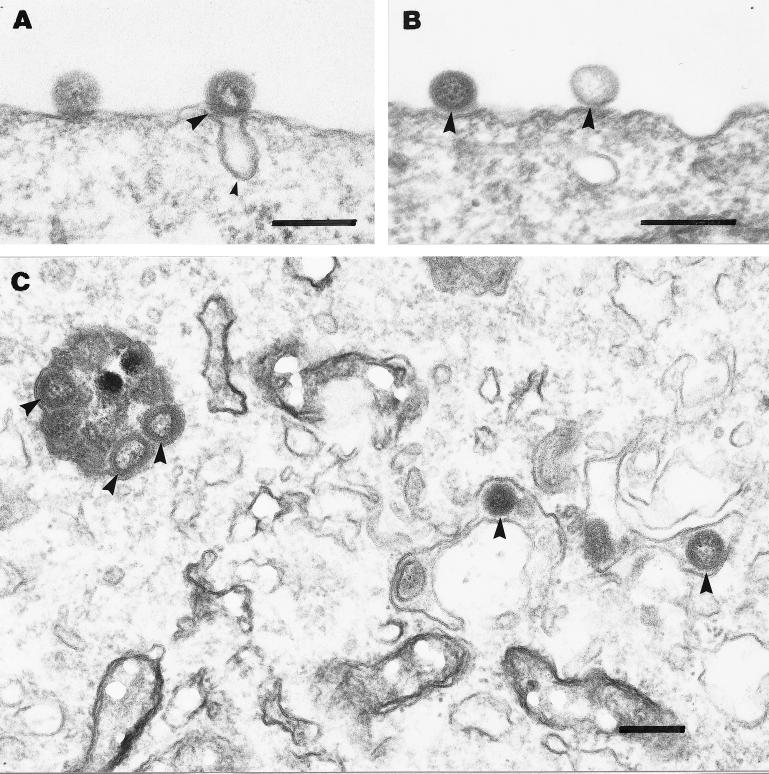
We have investigated the initial steps in the interaction between infectious salmon anemia virus (ISAV) and cultured cells from Atlantic salmon (SHK-1 cell line). Using radioactively or fluorescently labelled viral particles we have studied the binding and fusion kinetics and the effect of pH on binding, uptake, and fusion of ISAV to SHK-1 cells and liposomes. As pH in the medium was reduced from 7.5 to 4.5, the association of virus to the cells was nearly doubled. The same effect of pH was observed when fusion between ISAV and liposomes was analyzed. In addition, the binding of ISAV to intact SHK-1 cells and to cell membrane proteins blotted onto filters was neuraminidase sensitive. However, the increased binding induced by low pH was not neuraminidase sensitive, probably reflecting activation of a fusion peptide at low pH. By using confocal fluorescence microscopy, the increased fusion of fluorescently labelled ISAV with the plasma membrane due to low pH could be demonstrated. When vacuolar pH in the cells was raised during inoculation with chloroquine or ammonium chloride, both electron and confocal microscopy showed accumulation of ISAV in endosomes and lysosomes. Production of infectious virus could be increased by lowering the extracellular pH during infection. Furthermore, chloroquine present during virus inoculation also caused a reduction in the synthesis of viral proteins in ISAV-infected cells as well as in the production of infective virus. These results indicate that ISAV binds to sialic acid residues on the cell surface and that the fusion between virus and cell membrane takes place in the acid environment of endosomes. This provides further evidence for a high degree of similarity between ISAV and influenza virus and extends the basis for the classification of this virus as a member of the Orthomyxoviridae family.
Figures







References
-
- Berg T, Tolleshaug H. The effects of ammonium ions and chloroquine on uptake and degradation of 125I-labeled asialo-fetuin in isolated rat hepatocytes. Biochem Pharmacol. 1980;29:917–925. - PubMed
-
- Blumenthal R, Bali-Puri A, Walter A, Covell D, Eidelman O. pH-dependent fusion of vesicular stomatitis virus with Vero cells. Measurement by dequenching of octadecyl rhodamine fluorescence. J Biol Chem. 1987;262:13614–13619. . (Erratum, 263:588, 1988.) - PubMed
-
- Bullough P A, Hughson F M, Skehel J J, Wiley D C. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature. 1994;371:37–43. - PubMed
-
- Crider B P, Xie X S, Stone D K. Bafilomycin inhibits proton flow through the H+ channel of vacuolar proton pumps. J Biol Chem. 1994;269:17379–17381. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
