

Alleviation of human papillomavirus E2-mediated transcriptional repression via formation of a TATA binding protein (or TFIID)-TFIIB-RNA polymerase II-TFIIF preinitiation complex
- PMID: 10594014
- PMCID: PMC85067
- DOI: 10.1128/MCB.20.1.113-125.2000
Alleviation of human papillomavirus E2-mediated transcriptional repression via formation of a TATA binding protein (or TFIID)-TFIIB-RNA polymerase II-TFIIF preinitiation complex
Abstract
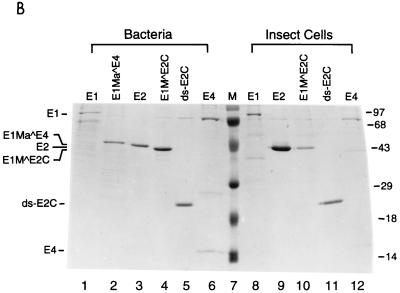
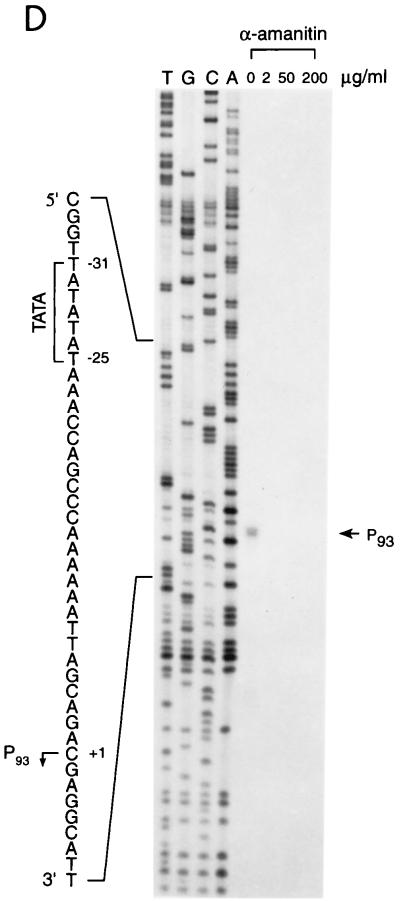
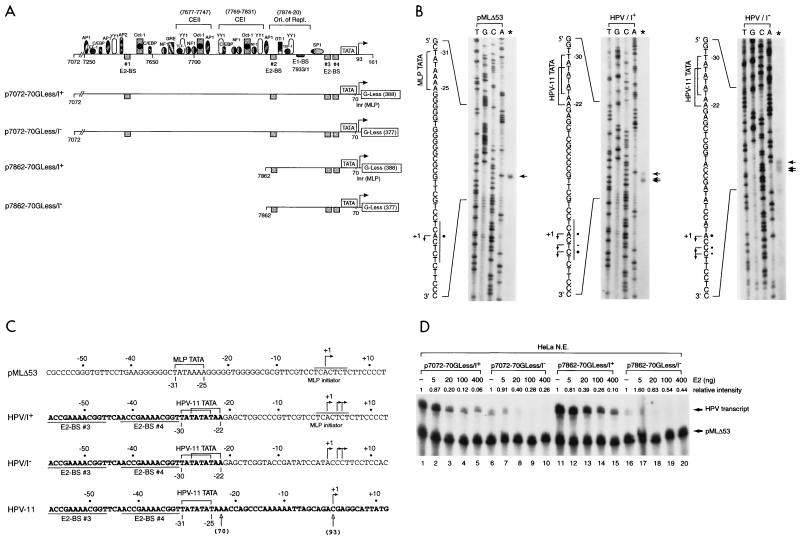
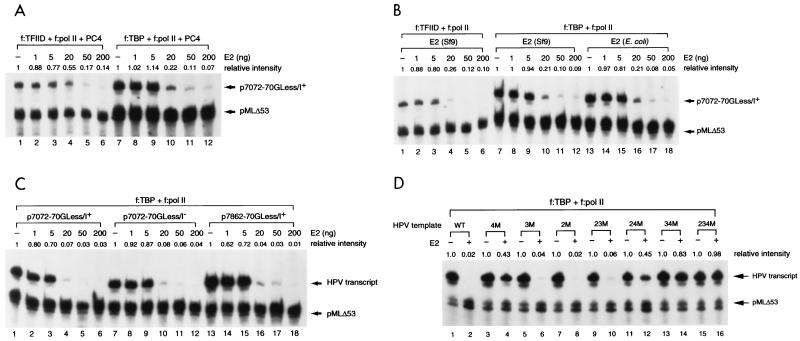
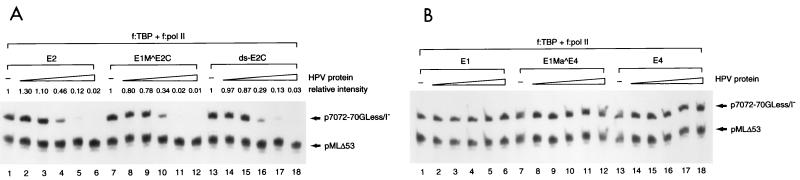
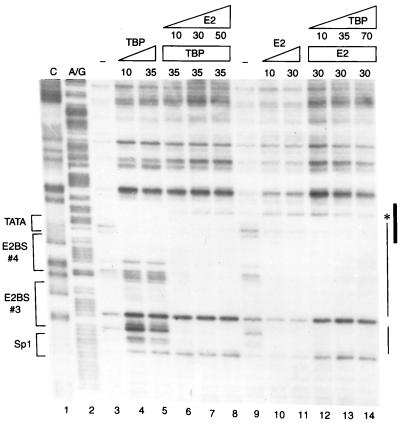
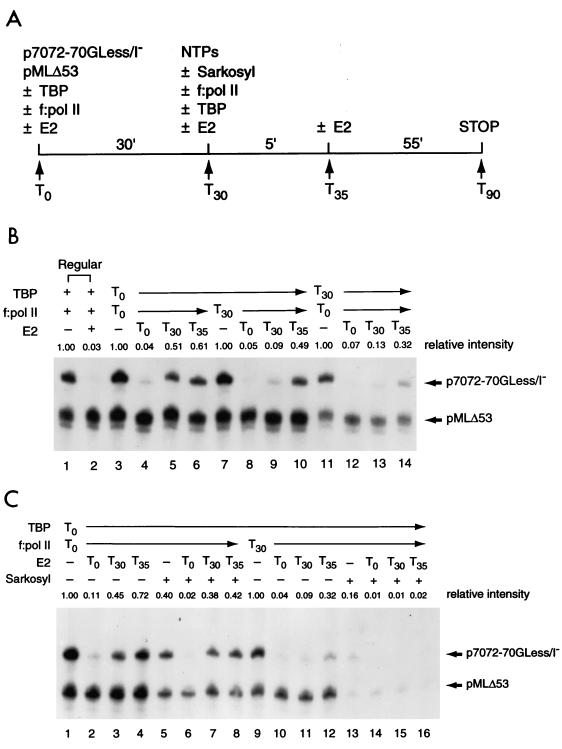
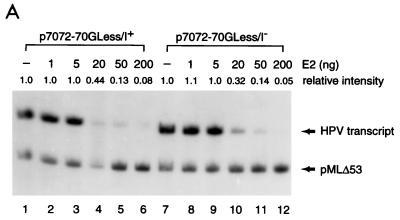
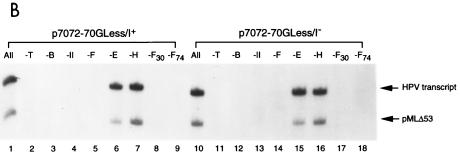
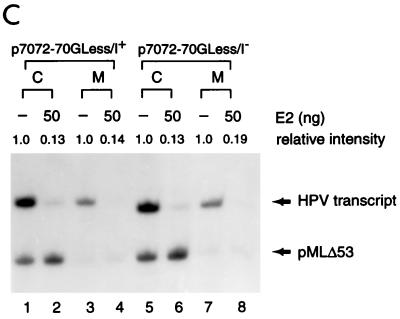
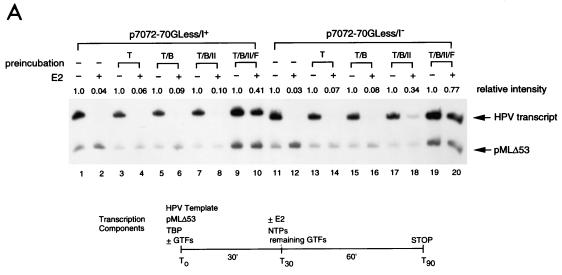
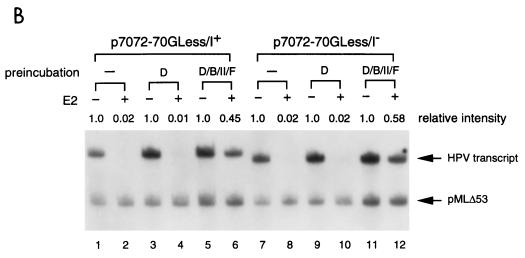
Transcription in human papillomaviruses (HPVs) is mainly regulated by cellular transcription factors and virus-encoded E2 proteins that act as sequence-specific DNA-binding proteins. Although the functions of E2 as a transcriptional activator and a repressor have been well documented, the role of cellular factors involved in E2-mediated regulation of the HPV promoters and the mechanism by which E2 modulates viral gene expression remain unclear. Using reconstituted cell-free transcription systems, we found that cellular enhancer-binding factors and general cofactors, such as TAF(II)s, TFIIA, Mediator, and PC4, are not required for E2-mediated repression. Unlike other transcriptional repressors that function through recruitment of histone deacetylase or corepressor complexes, HPV E2 is able to directly target components of the general transcription machinery to exert its repressor activity on the natural HPV E6 promoter. Interestingly, preincubation of TATA binding protein (TBP) or TFIID with HPV template is not sufficient to overcome E2-mediated repression, which can be alleviated only via formation of a minimal TBP (or TFIID)-TFIIB-RNA polymerase II-TFIIF preinitiation complex. Our data therefore indicate that E2 does not simply work by displacing TBP or TFIID from binding to the adjacent TATA box. Instead, E2 appears to function as an active repressor that directly inhibits HPV transcription at steps after TATA recognition by TBP or TFIID.
Figures


Similar articles
-
TATA-binding protein-associated factors enhance the recruitment of RNA polymerase II by transcriptional activators.J Biol Chem. 2001 Sep 7;276(36):34235-43. doi: 10.1074/jbc.M102463200. Epub 2001 Jul 16. J Biol Chem. 2001. PMID: 11457828
-
Bovine papillomavirus type 1 E2 transcriptional regulators directly bind two cellular transcription factors, TFIID and TFIIB.J Virol. 1995 Oct;69(10):6323-34. doi: 10.1128/JVI.69.10.6323-6334.1995. J Virol. 1995. PMID: 7666533 Free PMC article.
-
Intermediates in formation and activity of the RNA polymerase II preinitiation complex: holoenzyme recruitment and a postrecruitment role for the TATA box and TFIIB.Genes Dev. 1999 Jan 1;13(1):49-63. doi: 10.1101/gad.13.1.49. Genes Dev. 1999. PMID: 9887099 Free PMC article.
-
Mechanisms of transcriptional activation and repression can both involve TFIID.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):517-26. doi: 10.1098/rstb.1996.0050. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735274 Review.
-
The general transcription machinery and general cofactors.Crit Rev Biochem Mol Biol. 2006 May-Jun;41(3):105-78. doi: 10.1080/10409230600648736. Crit Rev Biochem Mol Biol. 2006. PMID: 16858867 Review.
Cited by
-
Interaction of papillomavirus E2 protein with the Brm chromatin remodeling complex leads to enhanced transcriptional activation.J Virol. 2007 Mar;81(5):2213-20. doi: 10.1128/JVI.01746-06. Epub 2006 Dec 6. J Virol. 2007. PMID: 17151122 Free PMC article.
-
Disruption of CTCF-YY1-dependent looping of the human papillomavirus genome activates differentiation-induced viral oncogene transcription.PLoS Biol. 2018 Oct 25;16(10):e2005752. doi: 10.1371/journal.pbio.2005752. eCollection 2018 Oct. PLoS Biol. 2018. PMID: 30359362 Free PMC article.
-
Human mediator enhances activator-facilitated recruitment of RNA polymerase II and promoter recognition by TATA-binding protein (TBP) independently of TBP-associated factors.Mol Cell Biol. 2003 Sep;23(17):6229-42. doi: 10.1128/MCB.23.17.6229-6242.2003. Mol Cell Biol. 2003. PMID: 12917344 Free PMC article.
-
Acetylation of E2 by P300 Mediates Topoisomerase Entry at the Papillomavirus Replicon.J Virol. 2019 Mar 21;93(7):e02224-18. doi: 10.1128/JVI.02224-18. Print 2019 Apr 1. J Virol. 2019. PMID: 30651357 Free PMC article.
-
Origin of hepatitis delta virus mRNA.J Virol. 2000 Aug;74(16):7204-10. doi: 10.1128/jvi.74.16.7204-7210.2000. J Virol. 2000. PMID: 10906174 Free PMC article.
References
-
- Alexander K A, Phelps W C. A fluorescence anisotropy study of DNA binding by HPV-11 E2C protein: a hierarchy of E2-binding sites. Biochemistry. 1996;35:9864–9872. - PubMed
-
- Auble D T, Hansen K E, Mueller C G F, Lane W S, Thorner J, Hahn S. Mot1, a global repressor of RNA polymerase II transcription, inhibits TBP binding to DNA by an ATP-dependent mechanism. Genes Dev. 1994;8:1920–1934. - PubMed
-
- Burley S K, Roeder R G. TATA box mimicry by TFIID: autoinhibition of pol II transcription. Cell. 1998;94:551–553. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources