

Establishment of distinct MyoD, E2A, and twist DNA binding specificities by different basic region-DNA conformations
- PMID: 10594029
- PMCID: PMC85082
- DOI: 10.1128/MCB.20.1.261-272.2000
Establishment of distinct MyoD, E2A, and twist DNA binding specificities by different basic region-DNA conformations
Abstract
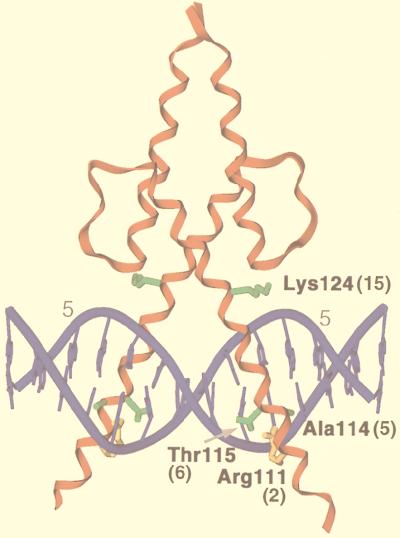
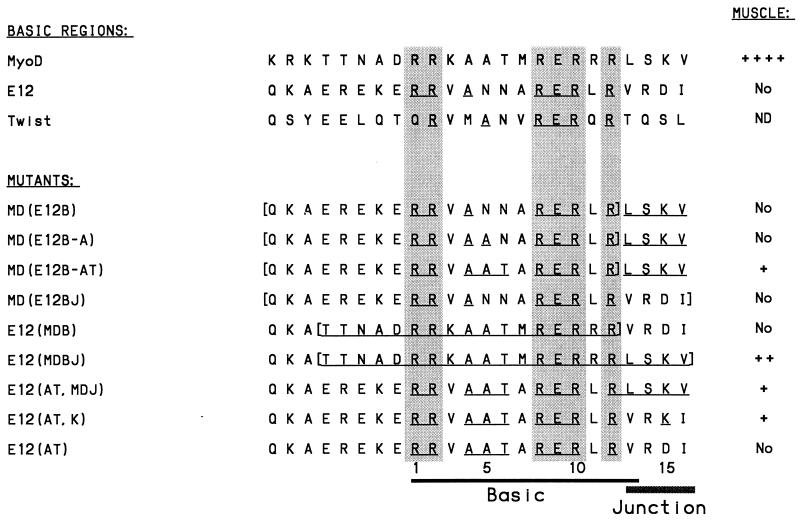
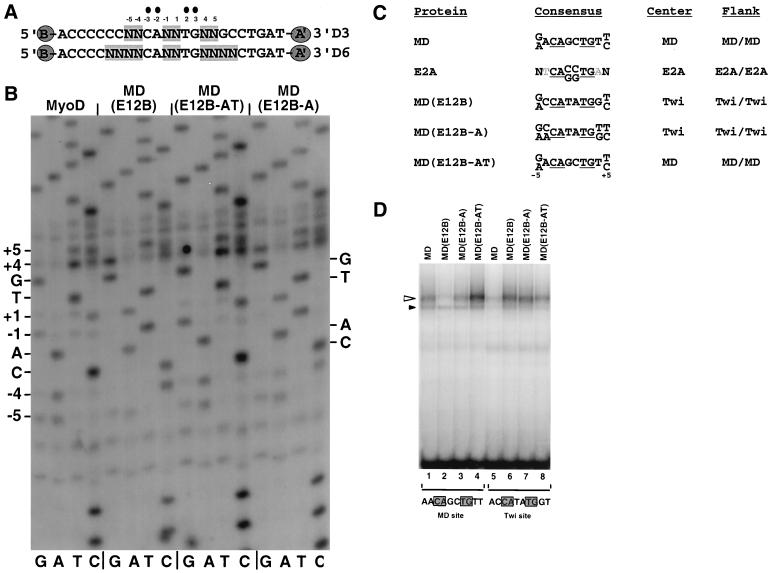
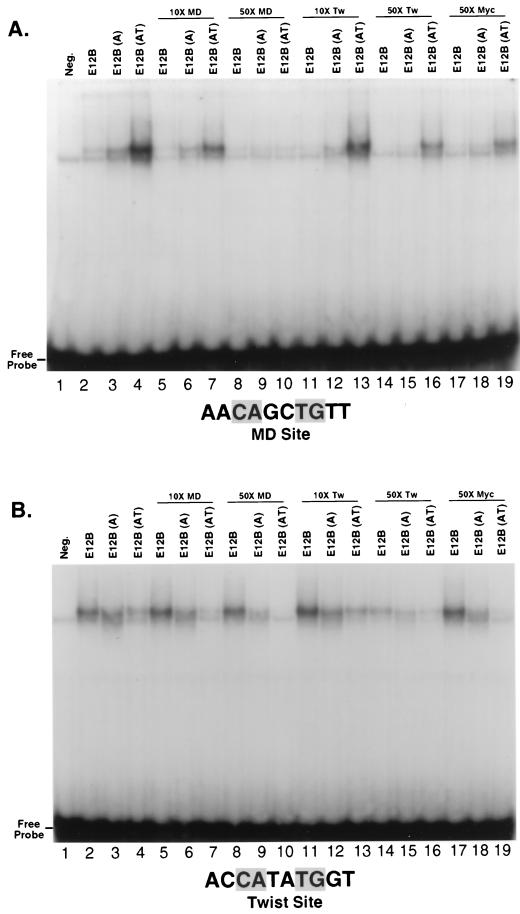
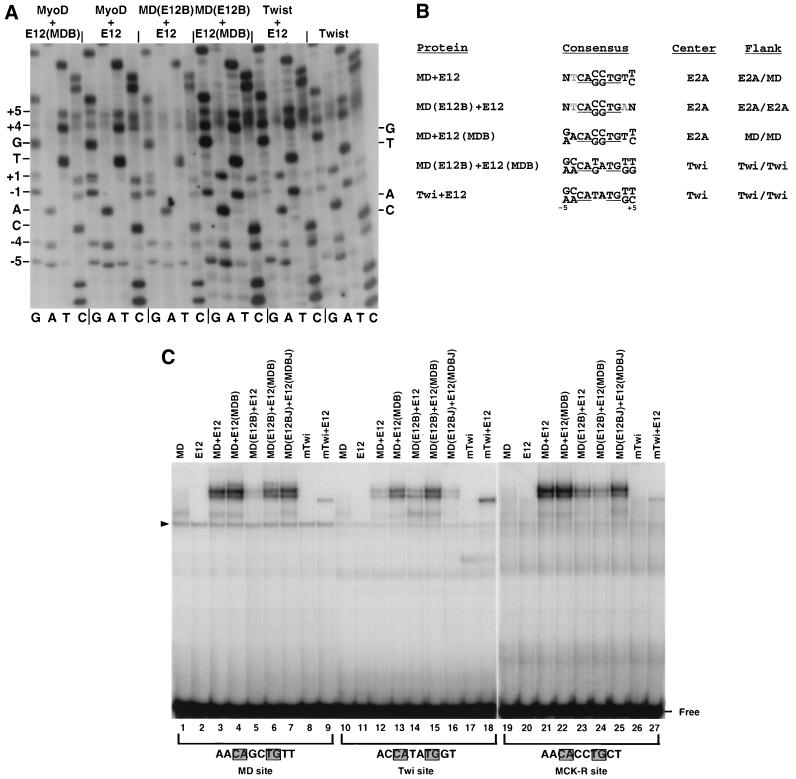
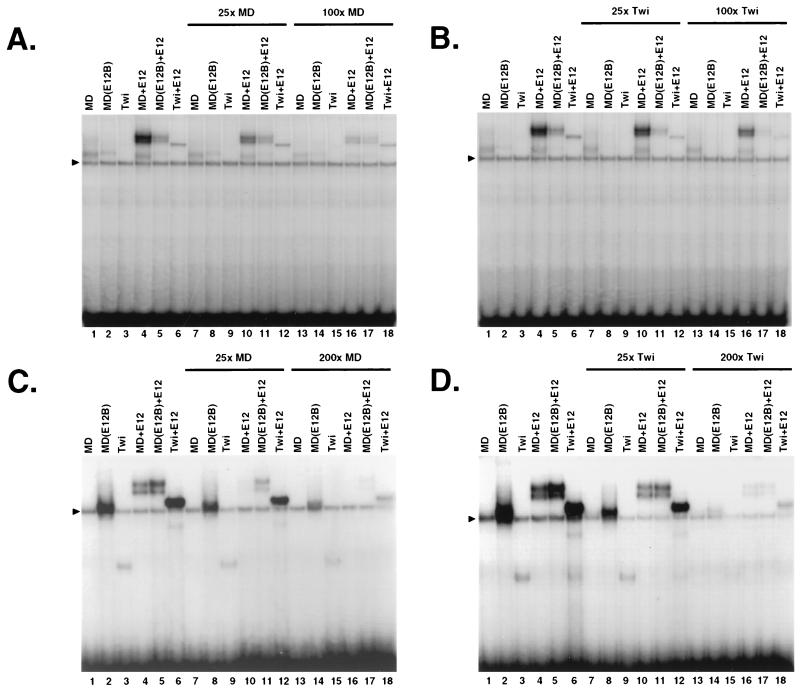
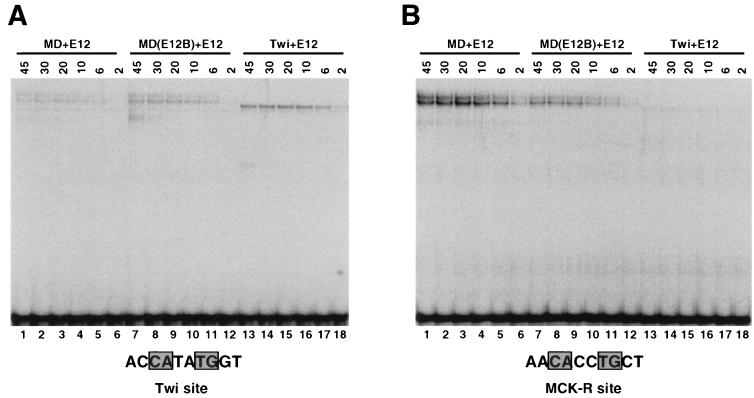
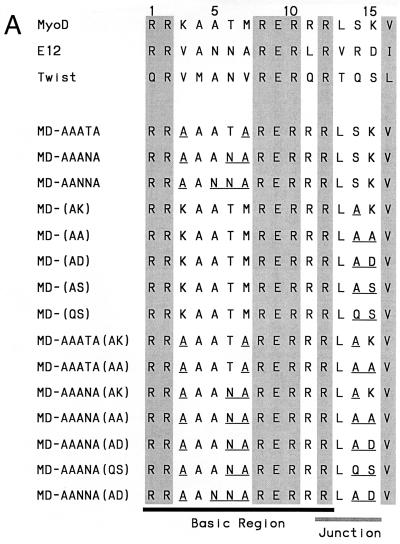
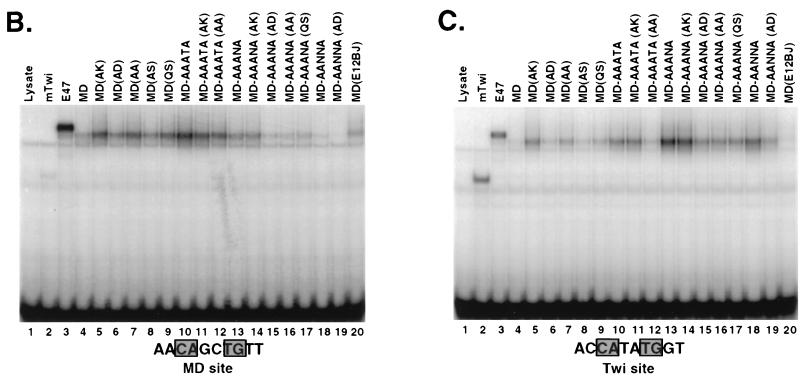
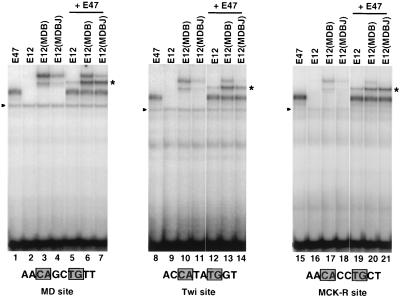
Basic helix-loop-helix (bHLH) proteins perform a wide variety of biological functions. Most bHLH proteins recognize the consensus DNA sequence CAN NTG (the E-box consensus sequence is underlined) but acquire further functional specificity by preferring distinct internal and flanking bases. In addition, induction of myogenesis by MyoD-related bHLH proteins depends on myogenic basic region (BR) and BR-HLH junction residues that are not essential for binding to a muscle-specific site, implying that their BRs may be involved in other critical interactions. We have investigated whether the myogenic residues influence DNA sequence recognition and how MyoD, Twist, and their E2A partner proteins prefer distinct CAN NTG sites. In MyoD, the myogenic BR residues establish specificity for particular CAN NTG sites indirectly, by influencing the conformation through which the BR helix binds DNA. An analysis of DNA binding by BR and junction mutants suggests that an appropriate BR-DNA conformation is necessary but not sufficient for myogenesis, supporting the model that additional interactions with this region are important. The sequence specificities of E2A and Twist proteins require the corresponding BR residues. In addition, mechanisms that position the BR allow E2A to prefer distinct half-sites as a heterodimer with MyoD or Twist, indicating that the E2A BR can be directed toward different targets by dimerization with different partners. Our findings indicate that E2A and its partner bHLH proteins bind to CAN NTG sites by adopting particular preferred BR-DNA conformations, from which they derive differences in sequence recognition that can be important for functional specificity.
Figures
Similar articles
-
The basic domain of myogenic basic helix-loop-helix (bHLH) proteins is the novel target for direct inhibition by another bHLH protein, Twist.Mol Cell Biol. 1997 Nov;17(11):6563-73. doi: 10.1128/MCB.17.11.6563. Mol Cell Biol. 1997. PMID: 9343420 Free PMC article.
-
DNA binding specificity of the basic-helix-loop-helix protein MASH-1.Biochemistry. 1995 Sep 5;34(35):11026-36. doi: 10.1021/bi00035a008. Biochemistry. 1995. PMID: 7669760
-
Acquisition of myogenic specificity by replacement of three amino acid residues from MyoD into E12.Science. 1992 May 15;256(5059):1027-30. doi: 10.1126/science.1317057. Science. 1992. PMID: 1317057
-
Muscle differentiation: more complexity to the network of myogenic regulators.Curr Opin Genet Dev. 1998 Oct;8(5):539-44. doi: 10.1016/s0959-437x(98)80008-7. Curr Opin Genet Dev. 1998. PMID: 9794824 Review.
-
Relationship between the structure and function of the transcriptional regulator E2A.J Biol Res (Thessalon). 2021 Jul 16;28(1):15. doi: 10.1186/s40709-021-00146-5. J Biol Res (Thessalon). 2021. PMID: 34271975 Free PMC article. Review.
Cited by
-
Cis-regulatory determinants of MyoD function.Nucleic Acids Res. 2018 Aug 21;46(14):7221-7235. doi: 10.1093/nar/gky388. Nucleic Acids Res. 2018. PMID: 30016497 Free PMC article.
-
The C. elegans Twist target gene, arg-1, is regulated by distinct E box promoter elements.Mech Dev. 2007 May;124(5):377-89. doi: 10.1016/j.mod.2007.01.005. Epub 2007 Feb 6. Mech Dev. 2007. PMID: 17369030 Free PMC article.
-
Mechanisms of Binding Specificity among bHLH Transcription Factors.Int J Mol Sci. 2021 Aug 24;22(17):9150. doi: 10.3390/ijms22179150. Int J Mol Sci. 2021. PMID: 34502060 Free PMC article. Review.
-
Computational modeling of the bHLH domain of the transcription factor TWIST1 and R118C, S144R and K145E mutants.BMC Bioinformatics. 2012 Jul 28;13:184. doi: 10.1186/1471-2105-13-184. BMC Bioinformatics. 2012. PMID: 22839202 Free PMC article.
-
Constitutive instability of muscle regulatory factor Myf5 is distinct from its mitosis-specific disappearance, which requires a D-box-like motif overlapping the basic domain.Mol Cell Biol. 2000 Dec;20(23):8923-32. doi: 10.1128/MCB.20.23.8923-8932.2000. Mol Cell Biol. 2000. PMID: 11073992 Free PMC article.
References
-
- Anthony-Cahill S J, Benfield P A, Fairman R, Wasserman Z R, Brenner S L, Stafford W F, Altenbach C, Hubbell W L, DeGrado W F. Molecular characterization of helix-loop-helix peptides. Science. 1992;255:979–983. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources