

The G-protein beta subunit GPB1 is required for mating and haploid fruiting in Cryptococcus neoformans
- PMID: 10594037
- PMCID: PMC85090
- DOI: 10.1128/MCB.20.1.352-362.2000
The G-protein beta subunit GPB1 is required for mating and haploid fruiting in Cryptococcus neoformans
Abstract
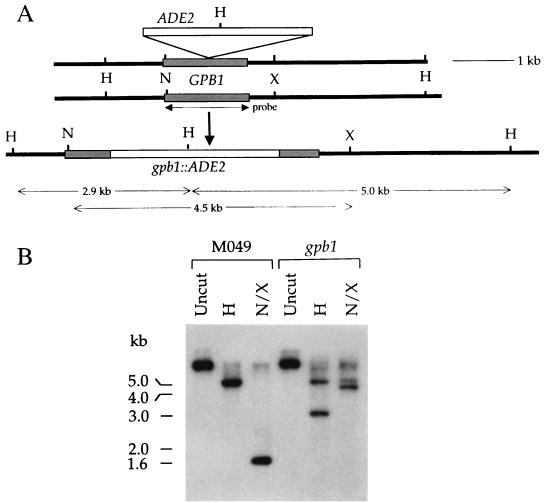
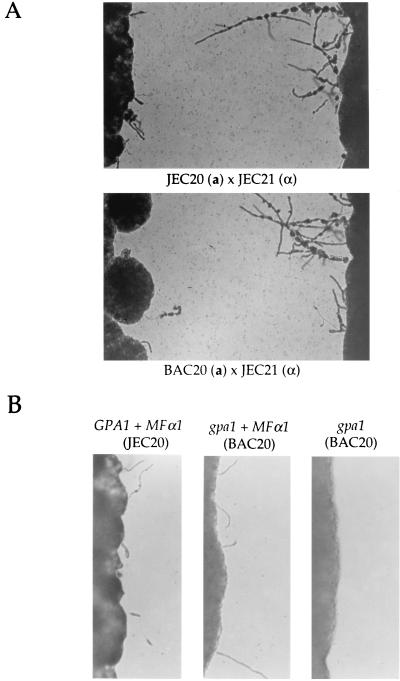
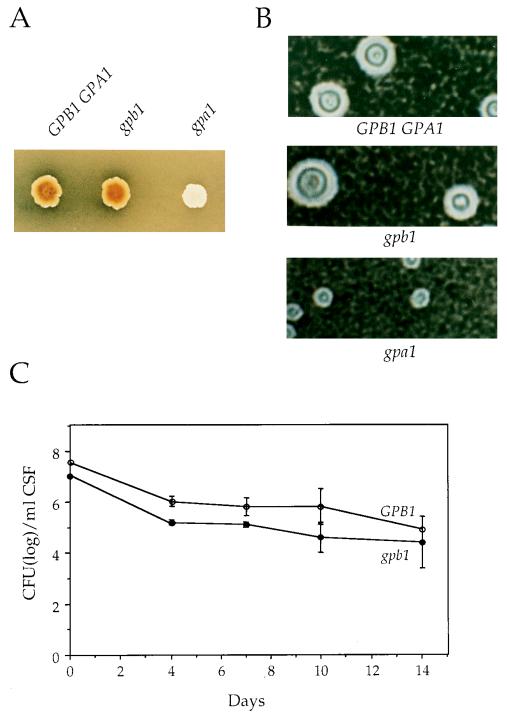
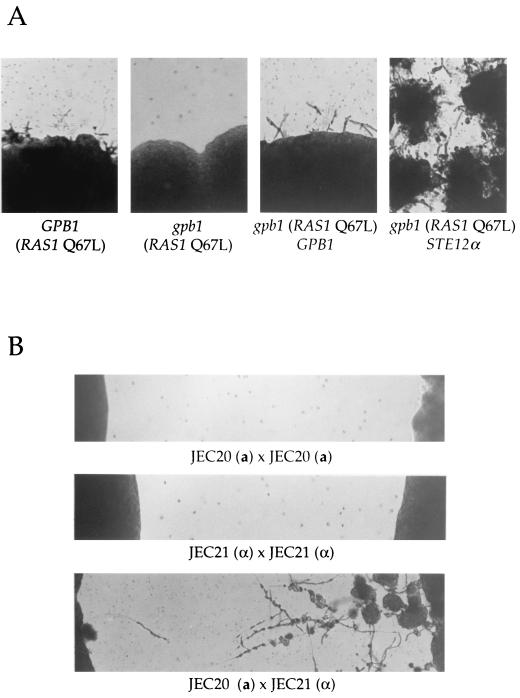
Cryptococcus neoformans is an opportunistic fungal pathogen with a defined sexual cycle. The gene encoding a heterotrimeric G-protein beta subunit, GPB1, was cloned and disrupted. gpb1 mutant strains are sterile, indicating a role for this gene in mating. GPB1 plays an active role in mediating responses to pheromones in early mating steps (conjugation tube formation and cell fusion) and signals via a mitogen-activated protein (MAP) kinase cascade in both MATalpha and MATa cells. The functions of GPB1 are distinct from those of the Galpha protein GPA1, which functions in a nutrient-sensing cyclic AMP (cAMP) pathway required for mating, virulence factor induction, and virulence. gpb1 mutant strains are also defective in monokaryotic fruiting in response to nitrogen starvation. We show that MATa cells stimulate monokaryotic fruiting of MATalpha cells, possibly in response to mating pheromone, which may serve to disperse cells and spores to locate mating partners. In summary, the Gbeta subunit GPB1 and the Galpha subunit GPA1 function in distinct signaling pathways: one (GPB1) senses pheromones and regulates mating and haploid fruiting via a MAP kinase cascade, and the other (GPA1) senses nutrients and regulates mating, virulence factors, and pathogenicity via a cAMP cascade.
Figures



References
-
- Alspaugh, J. A., R. C. Davidson, and J. Heitman. Morphogenesis of Cryptococcus neoformans. In J. F. Ernst and A. Schmidt (ed.), Dimorphism in human pathogenic and apathogenic yeasts, in press. S. Karger, Basel, Switzerland.
-
- Clapham D E, Neer E J. New roles for G-protein βγ-dimers in transmembrane signalling. Nature. 1993;365:403–406. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases