

The general anesthetic propofol slows deactivation and desensitization of GABA(A) receptors
- PMID: 10594047
- PMCID: PMC6784967
- DOI: 10.1523/JNEUROSCI.19-24-10635.1999
The general anesthetic propofol slows deactivation and desensitization of GABA(A) receptors
Abstract
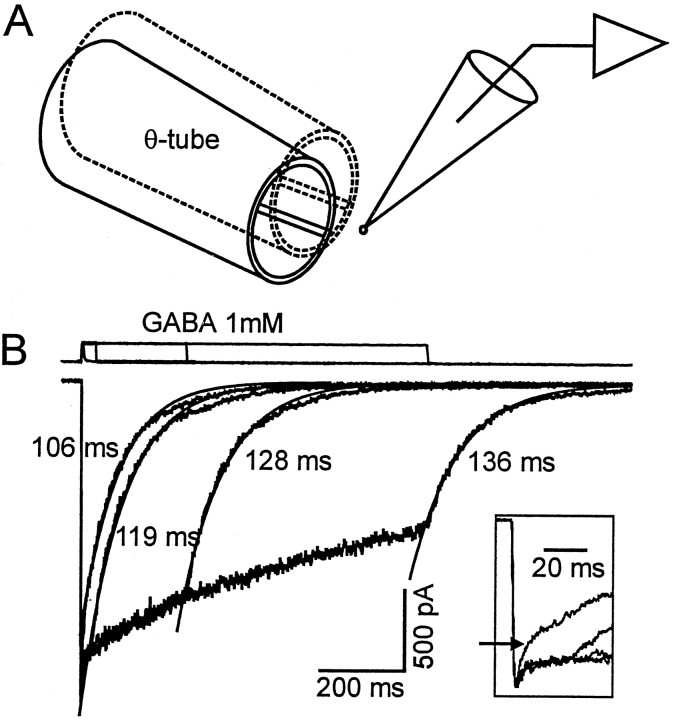
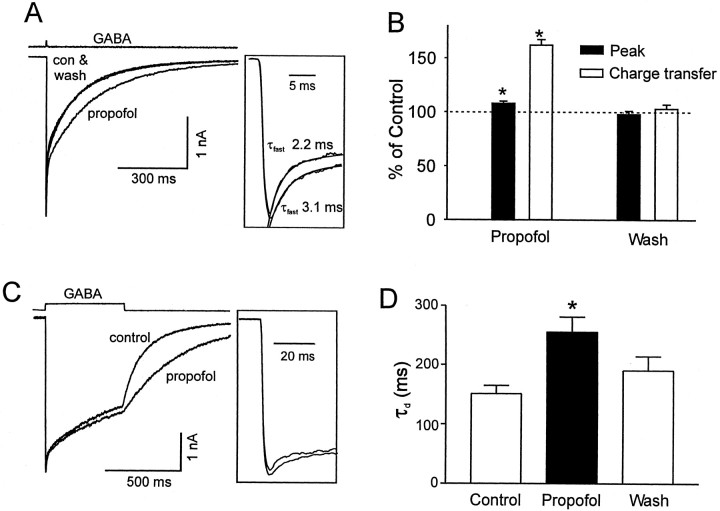
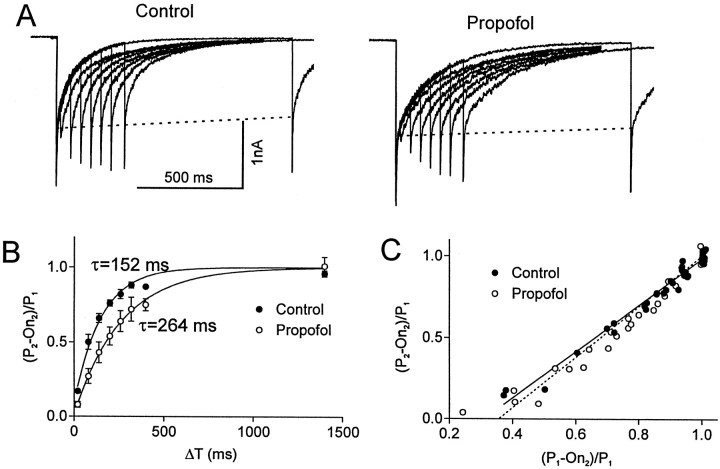
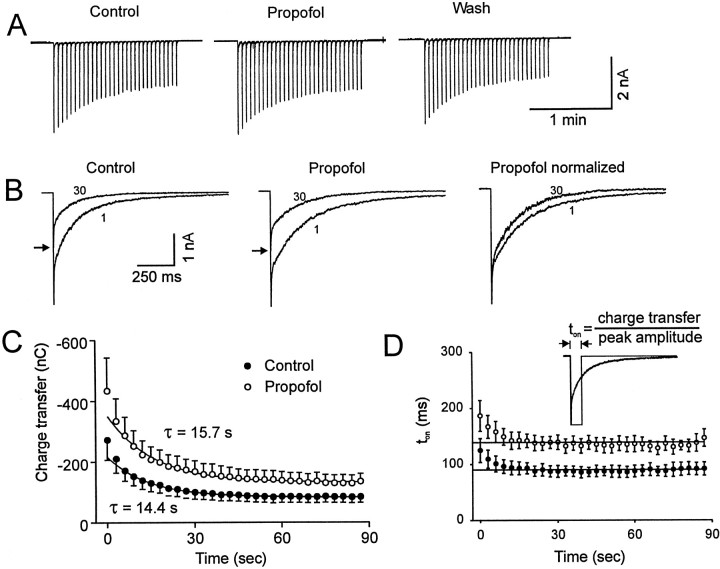
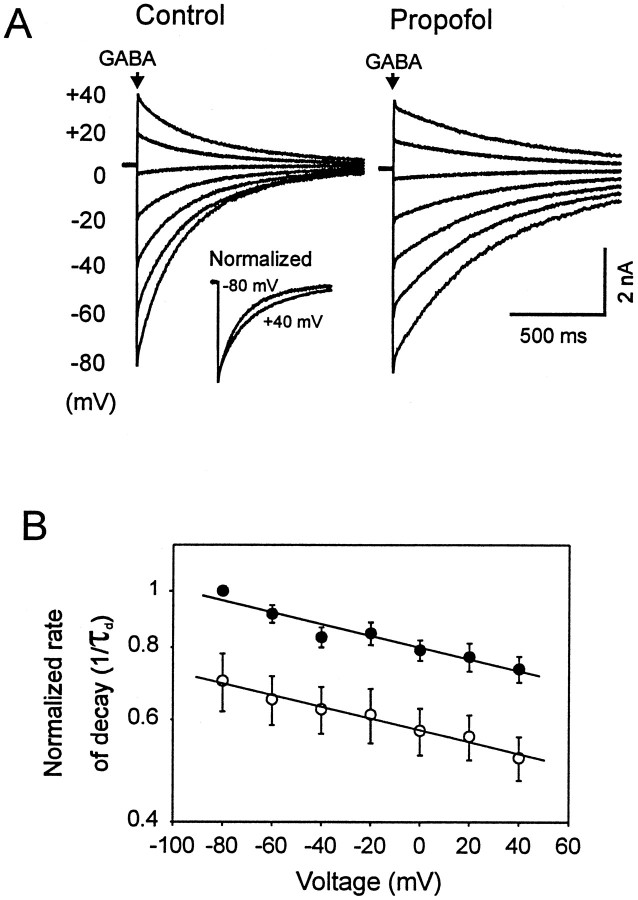
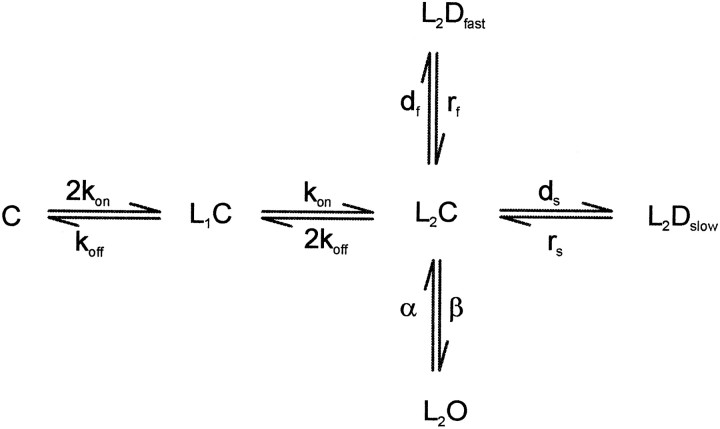
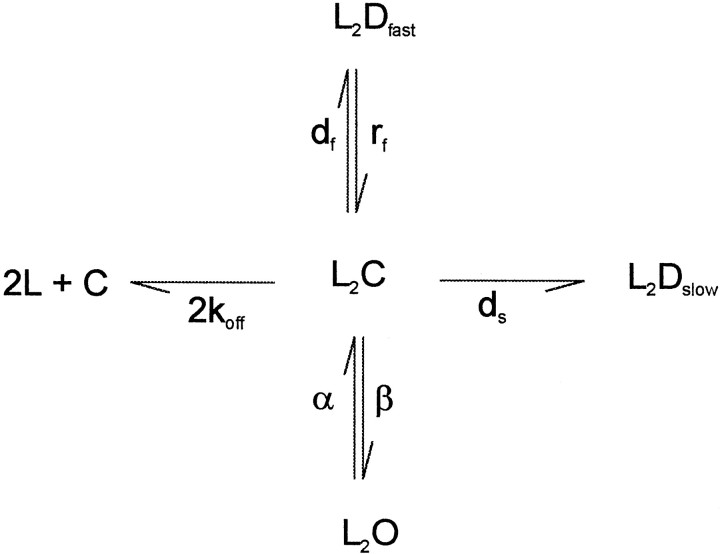
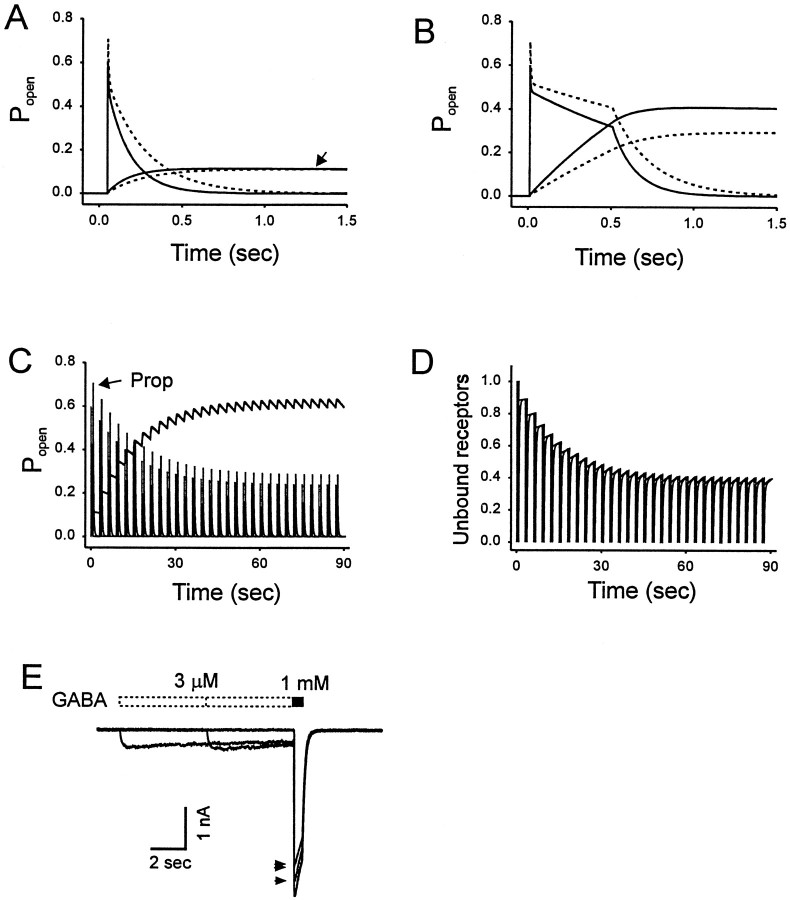
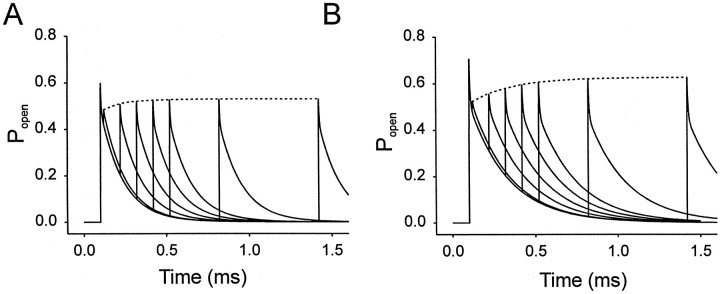
Propofol (2,6-di-isopropylphenol) has multiple actions on GABA(A) receptor function that act in concert to potentiate GABA-evoked currents. To understand how propofol influences inhibitory IPSCs, we examined the effects of propofol on responses to brief applications of saturating concentrations of GABA (1-30 mM). GABA was applied using a fast perfusion system to nucleated patches excised from hippocampal neurons. In this preparation, propofol (10 microM) had no detectable agonist effect but slowed the decay, increased the charge transfer (62%), and enhanced the peak amplitude (8%) of currents induced by brief pulses (3 msec) of GABA. Longer pulses (500 msec) of GABA induced responses that desensitized with fast (tau(f) = 1.5-4.5 msec) and slow (tau(s) = 1-3 sec) components and, after the removal of GABA, deactivated exponentially (tau(d) = 151 msec). Propofol prolonged this deactivation (tau(d) = 255 msec) and reduced the development of both fast and slow desensitization. Recovery from fast desensitization, assessed using pairs of brief pulses of GABA, paralleled the time course of deactivation, indicating that fast desensitization traps GABA on the receptor. With repetitive applications of pulses of GABA (0.33 Hz), the charge transfer per pulse declined exponentially (tau approximately 15 sec) to a steady-state value equal to approximately 40% of the initial response. Despite the increased charge transfer per pulse with propofol, the time course of the decline was unchanged. These experimental data were interpreted using computer simulations and a kinetic model that assumed fast and slow desensitization, as well as channel opening developed in parallel from a pre-open state. Our results suggest that propofol stabilizes the doubly liganded pre-open state without affecting the isomerization rate constants to and from the open state. Also, the rate constants for agonist dissociation and entry into the fast and slow desensitization states were reduced by propofol. The recovery rate constant from fast desensitization was slowed, whereas that from slow desensitization appeared to be unchanged. Taken together, the effects of propofol on GABA(A) receptors enhance channel opening, particularly under conditions that promote desensitization.
Figures
References
-
- Bai D, MacDonald JF, Orser BA. Midazolam and propofol modulation of tonic GABAergic current and transient IPSCs in cultured hippocampal neurons. Soc Neurosci Abstr. 1998;24:1832.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources