

Prominent axonopathy in the brain and spinal cord of transgenic mice overexpressing four-repeat human tau protein
- PMID: 10595944
- PMCID: PMC1866931
- DOI: 10.1016/S0002-9440(10)65533-2
Prominent axonopathy in the brain and spinal cord of transgenic mice overexpressing four-repeat human tau protein
Abstract
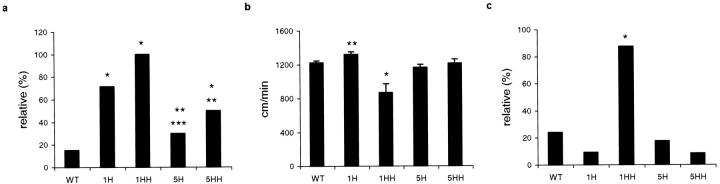
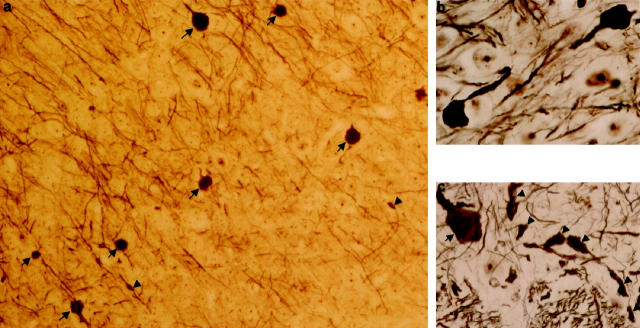
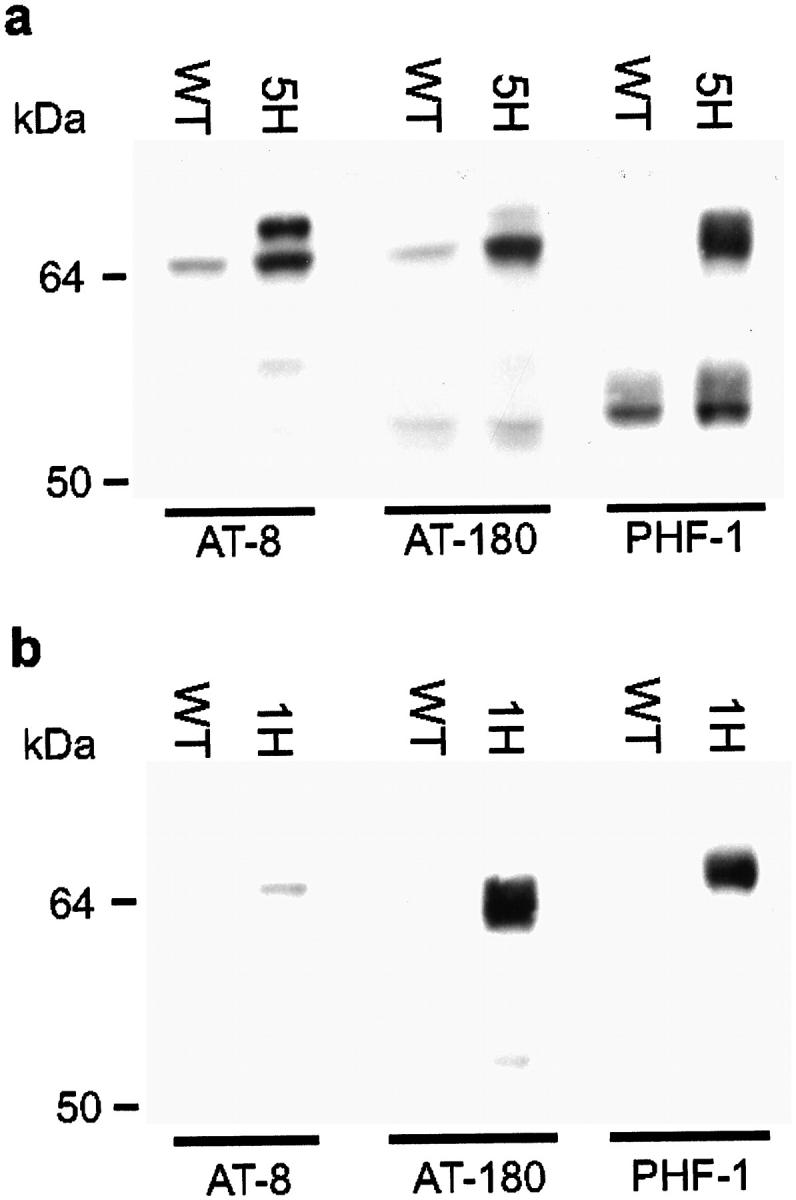
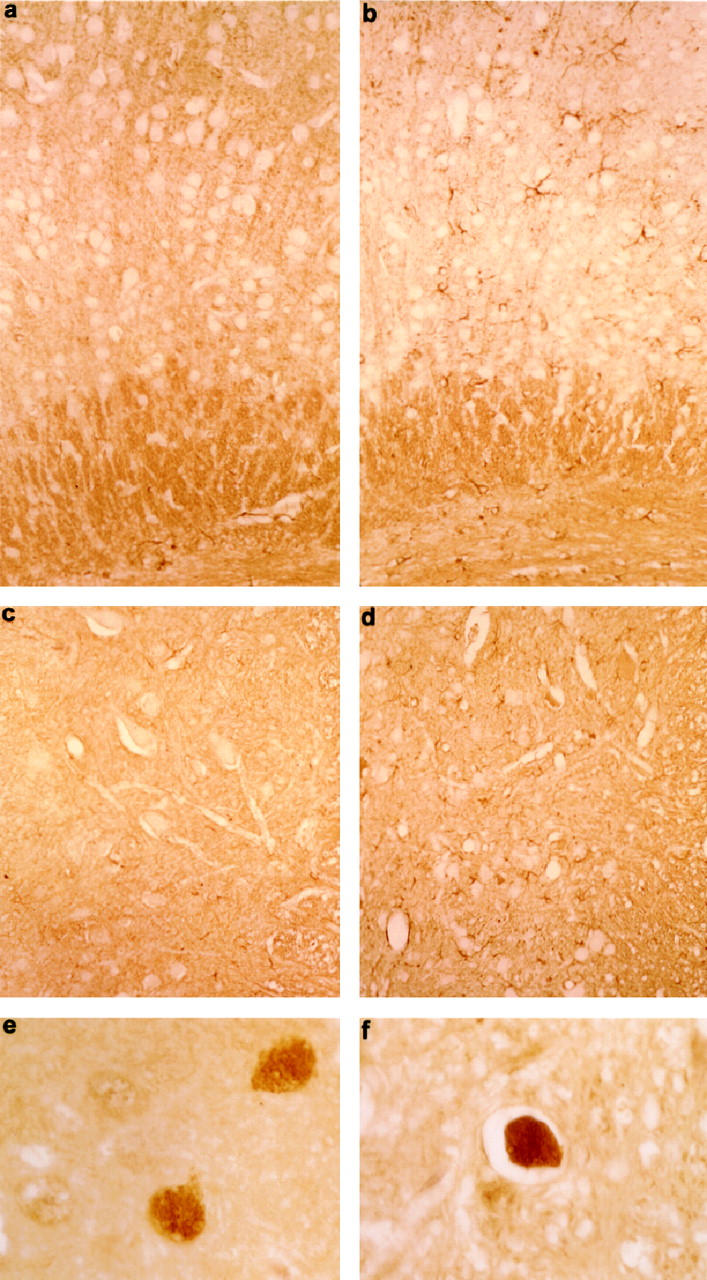
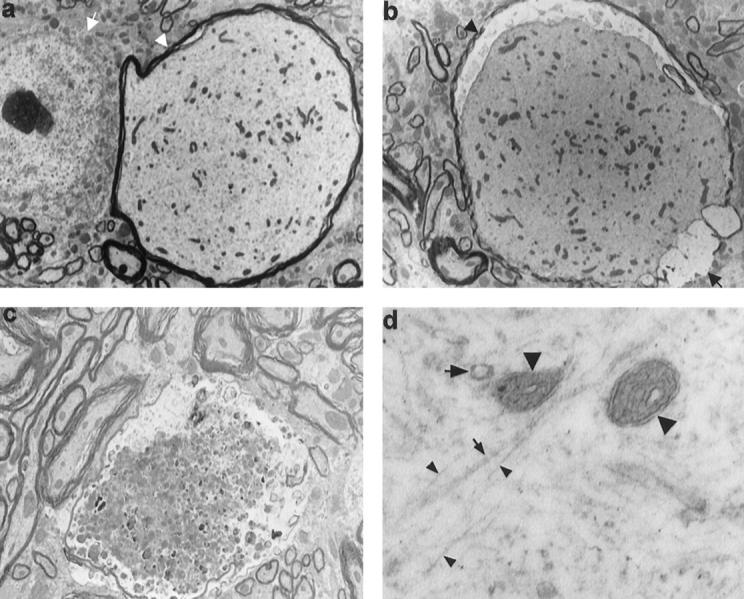
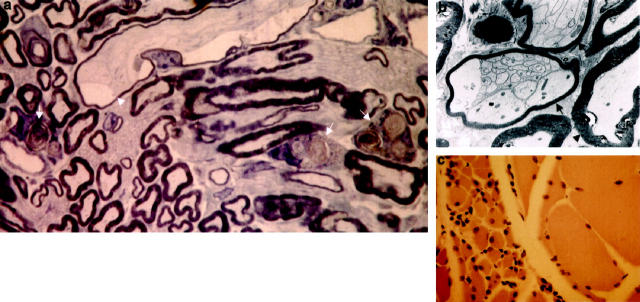
Mutations in the human tau gene cause frontotemporal dementia and parkinsonism linked to chromosome 17. Some mutations, including mutations in intron 10, induce increased levels of the functionally normal four-repeat tau protein isoform, leading to neurodegeneration. We generated transgenic mice that overexpress the four-repeat human tau protein isoform specifically in neurons. The transgenic mice developed axonal degeneration in brain and spinal cord. In the model, axonal dilations with accumulation of neurofilaments, mitochondria, and vesicles were documented. The axonopathy and the accompanying dysfunctional sensorimotor capacities were transgene-dosage related. These findings proved that merely increasing the concentration of the four-repeat tau protein isoform is sufficient to injure neurons in the central nervous system, without formation of intraneuronal neurofibrillary tangles. Evidence for astrogliosis and ubiquitination of accumulated proteins in the dilated part of the axon supported this conclusion. This transgenic model, overexpressing the longest isoform of human tau protein, recapitulates features of known neurodegenerative diseases, including Alzheimer's disease and other tauopathies. The model makes it possible to study the interaction with additional factors, to be incorporated genetically, or with other biological triggers that are implicated in neurodegeneration.
Figures



Similar articles
-
Prominent axonopathy and disruption of axonal transport in transgenic mice expressing human apolipoprotein E4 in neurons of brain and spinal cord.Am J Pathol. 2000 Nov;157(5):1495-510. doi: 10.1016/S0002-9440(10)64788-8. Am J Pathol. 2000. PMID: 11073810 Free PMC article.
-
Early axonopathy preceding neurofibrillary tangles in mutant tau transgenic mice.Am J Pathol. 2007 Sep;171(3):976-92. doi: 10.2353/ajpath.2007.070345. Epub 2007 Aug 9. Am J Pathol. 2007. PMID: 17690183 Free PMC article.
-
Attenuated neurodegenerative disease phenotype in tau transgenic mouse lacking neurofilaments.J Neurosci. 2001 Aug 15;21(16):6026-35. doi: 10.1523/JNEUROSCI.21-16-06026.2001. J Neurosci. 2001. PMID: 11487626 Free PMC article.
-
Transgenic mouse models of tauopathies: prospects for animal models of Pick's disease.Neurology. 2001 Jun;56(11 Suppl 4):S26-30. doi: 10.1212/wnl.56.suppl_4.s26. Neurology. 2001. PMID: 11402147 Review.
-
Tau and axonopathy in neurodegenerative disorders.Neuromolecular Med. 2002;2(2):131-50. doi: 10.1385/NMM:2:2:131. Neuromolecular Med. 2002. PMID: 12428808 Review.
Cited by
-
What Renders TAU Toxic.Front Neurol. 2013 Jun 10;4:72. doi: 10.3389/fneur.2013.00072. eCollection 2013. Front Neurol. 2013. PMID: 23772223 Free PMC article.
-
Multiple mechanisms of extracellular tau spreading in a non-transgenic tauopathy model.Am J Neurodegener Dis. 2012;1(3):316-33. Epub 2012 Nov 25. Am J Neurodegener Dis. 2012. PMID: 23383401 Free PMC article.
-
What we can learn from animal models about cerebral multi-morbidity.Alzheimers Res Ther. 2015 Jan 29;7(1):11. doi: 10.1186/s13195-015-0097-2. eCollection 2015. Alzheimers Res Ther. 2015. PMID: 25810783 Free PMC article.
-
The genetics of axonal transport and axonal transport disorders.PLoS Genet. 2006 Sep 29;2(9):e124. doi: 10.1371/journal.pgen.0020124. PLoS Genet. 2006. PMID: 17009871 Free PMC article. Review.
-
Prominent axonopathy and disruption of axonal transport in transgenic mice expressing human apolipoprotein E4 in neurons of brain and spinal cord.Am J Pathol. 2000 Nov;157(5):1495-510. doi: 10.1016/S0002-9440(10)64788-8. Am J Pathol. 2000. PMID: 11073810 Free PMC article.
References
-
- Wiche G, Oberkanins C, Himmler A: Molecular structure and function of microtubule-associated proteins. Int Rev Cytol 1991, 124:217-273 - PubMed
-
- Mandelkow EM, Mandelkow E: Tau in Alzheimer’s disease. Trends Cell Biol 1998, 8:125-127 - PubMed
-
- Andreadis A, Brown WM, Kosik KS: Structure and novel exons of the human tau gene. Biochemistry 1992, 31:10626-10633 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
