

The homeodomain protein vax1 is required for axon guidance and major tract formation in the developing forebrain
- PMID: 10601035
- PMCID: PMC317177
- DOI: 10.1101/gad.13.23.3092
The homeodomain protein vax1 is required for axon guidance and major tract formation in the developing forebrain
Abstract
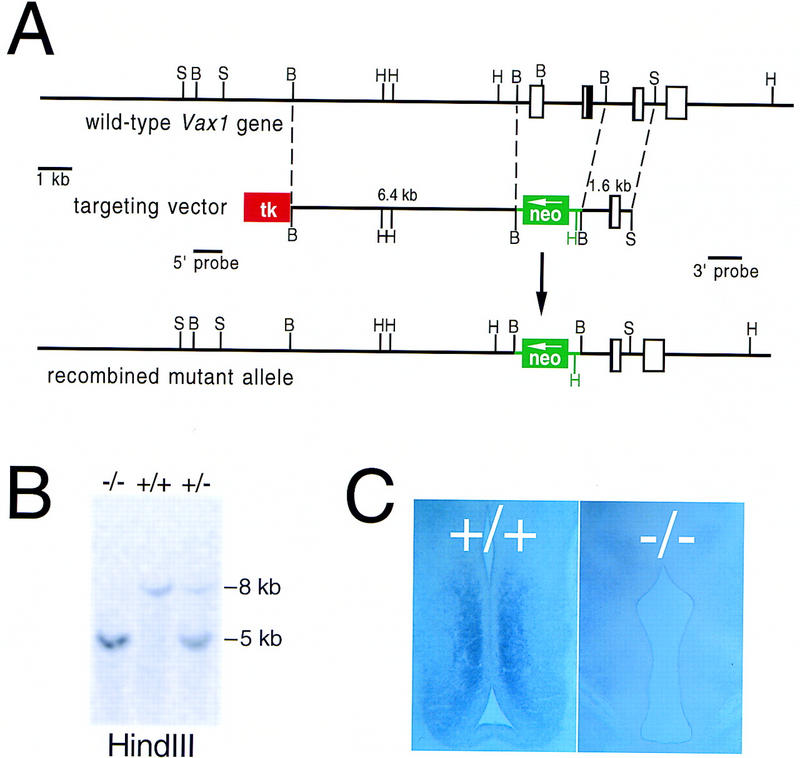
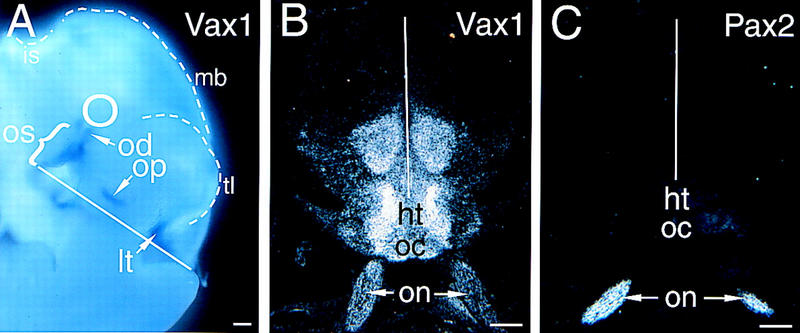
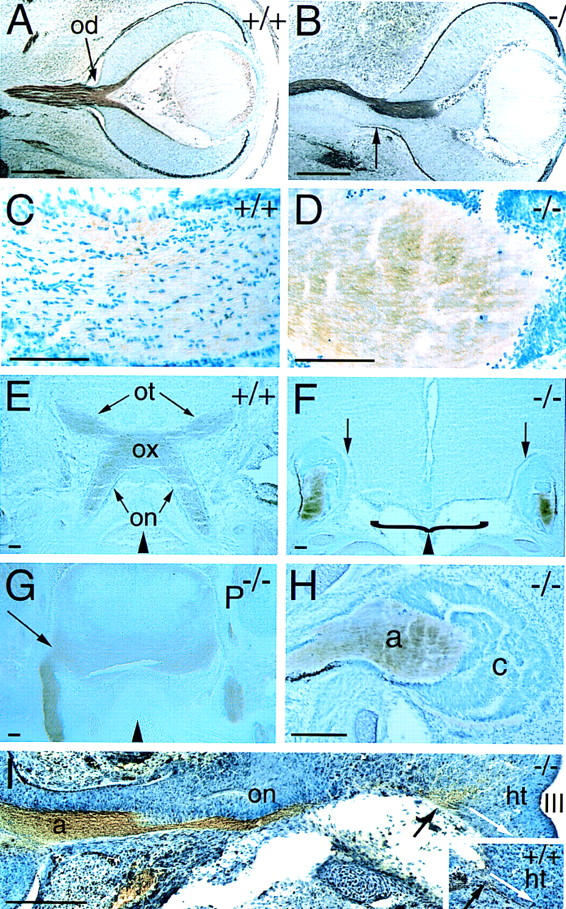
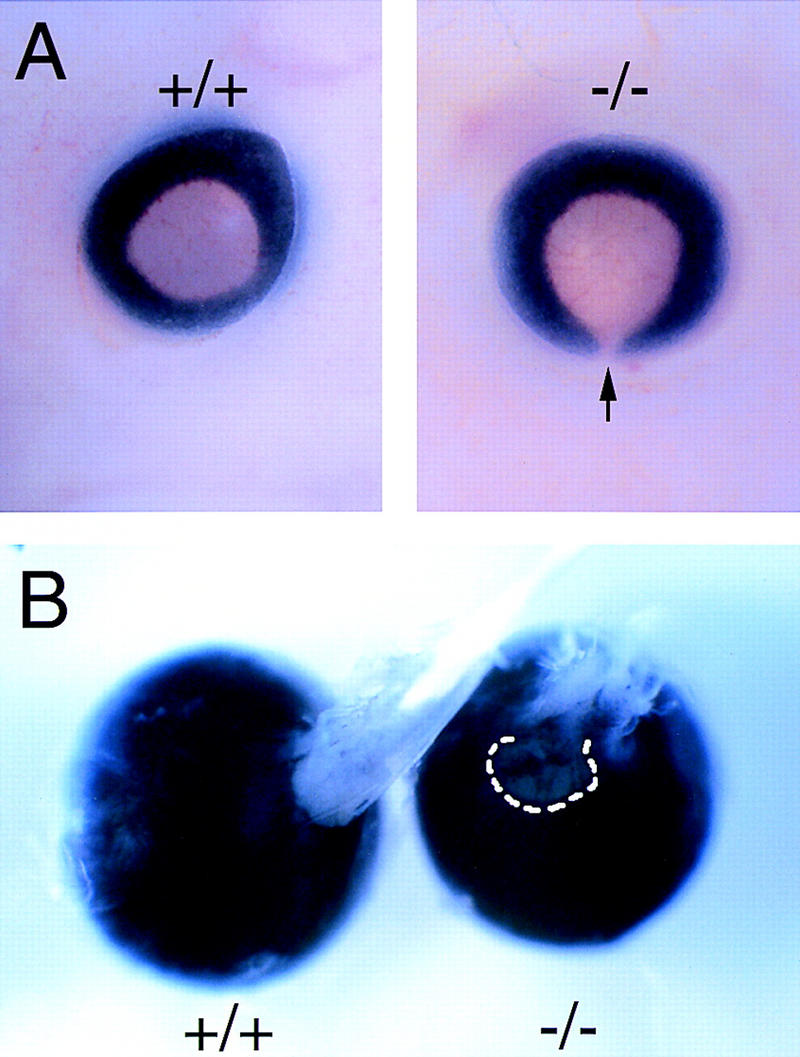
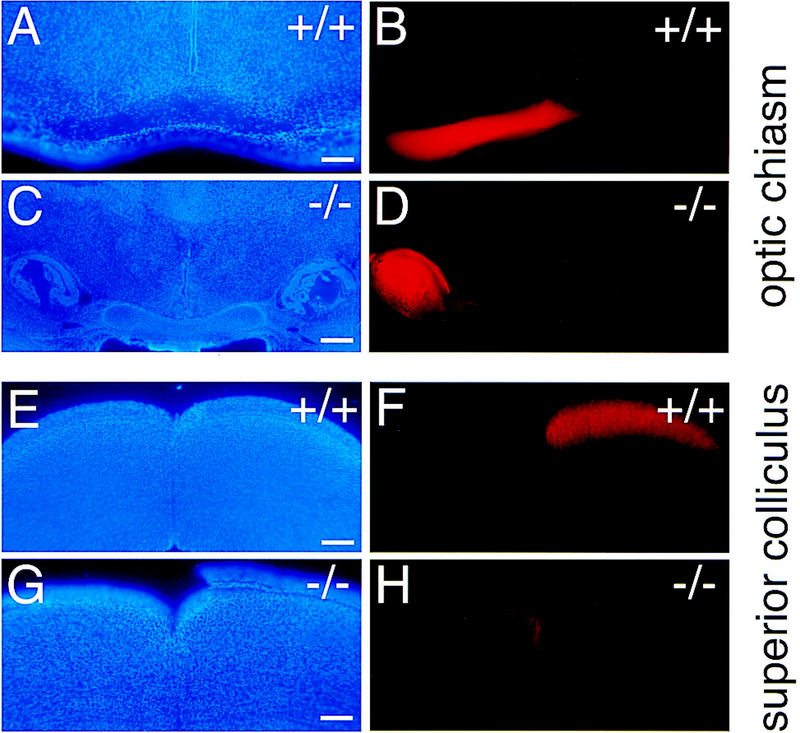
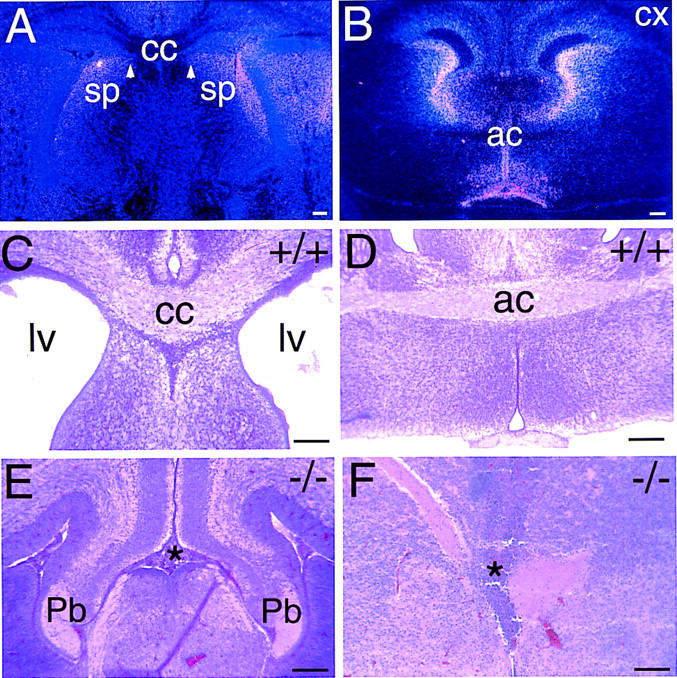
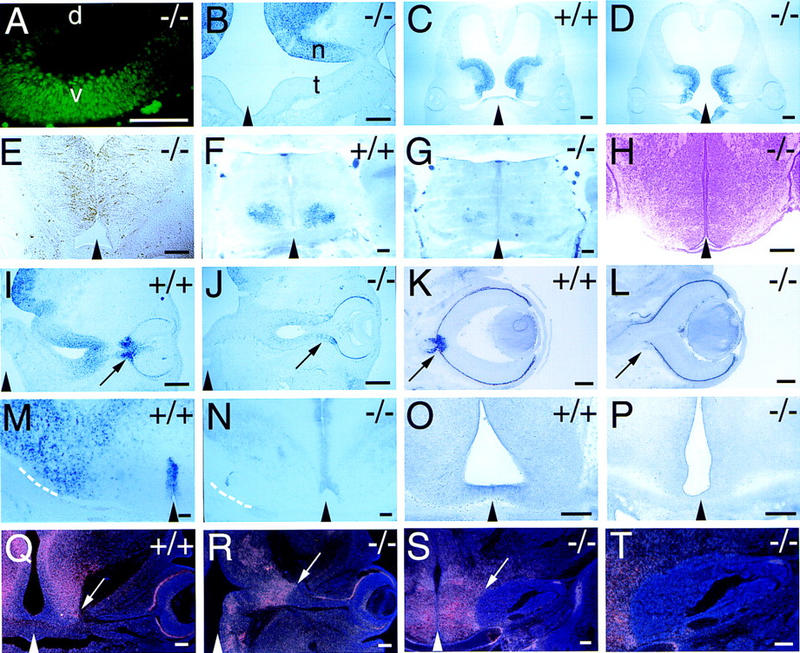
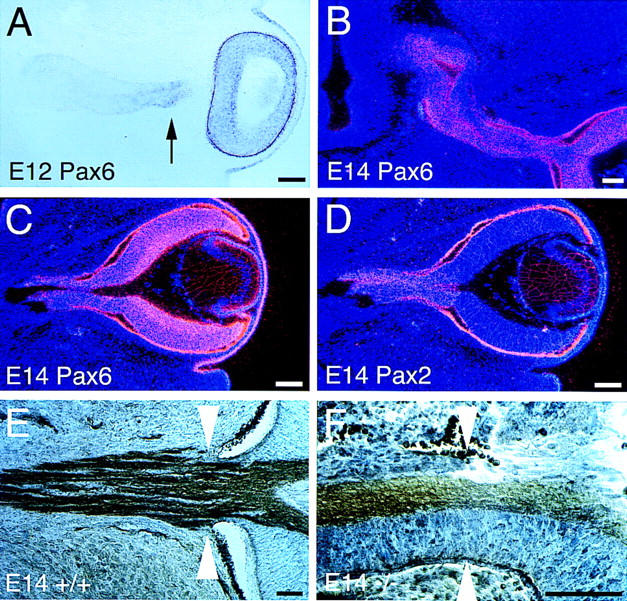
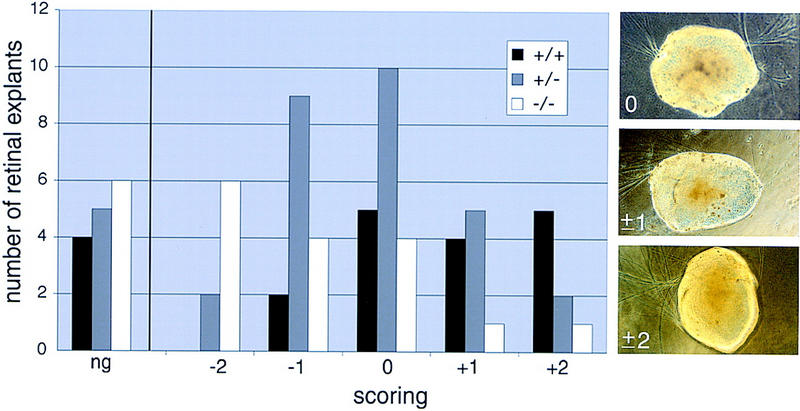
The homeodomain protein Vax1 is expressed in a highly circumscribed set of cells at the ventral anterior midline of the embryonic CNS. These cells populate the choroid fissure of the optic disk, the body of the optic stalk and nerve, the optic chiasm and ventral diencephalon, and the anterior midline zones that abut developing commissural tracts. We have generated mutant mice that lack Vax1. In these mice (1) the optic disks fail to close, leading to coloboma and loss of the eye-nerve boundary; (2) optic nerve glia fail to associate with and appear to repulse ingrowing retinal axons, resulting in a fascicle of axons that are completely segregated from optic nerve astrocytes; (3) retinal axons fail to penetrate the brain in significant numbers and fail to form an optic chiasm; and (4) axons in multiple commissural tracts of the anterior CNS, including the corpus callosum and the hippocampal and anterior commissures, fail to cross the midline. These axon guidance defects do not result from the death of normally Vax1(+) midline cells but, instead, correlate with markedly diminished expression of attractive guidance cues in these cells. Vax1 therefore regulates the guidance properties of a set of anterior midline cells that orchestrate axon trajectories in the developing mammalian forebrain.
Figures
References
-
- Anderson MG, Perkins GL, Chittick P, Shrigley RJ, Johnson WA. drifter, a Drosophila POU-domain transcription factor, is required for correct differentiation and migration of tracheal cells and midline glia. Genes & Dev. 1995;9:123–137. - PubMed
-
- Bartsch U, Kirchhoff F, Schachner M. Immunohistological localization of the adhesion molecules L1, N-CAM, and MAG in the developing and adult optic nerve of mice. J Comp Neurol. 1989;284:451–462. - PubMed
-
- Battye R, Stevens A, Jacobs JR. Axon repulsion from the midline of the Drosophila CNS requires slit function. Development. 1999;126:2475–2481. - PubMed
-
- Braisted JE, McLaughlin T, Wang HU, Friedman GC, Anderson DJ, O'Leary DDM. Graded and lamina-specific distributions of ligands of EphB receptor tyrosine kinases in the developing retinotectal system. Dev Biol. 1997;191:14–28. - PubMed
-
- Brose K, Bland KS, Wang KH, Arnott D, Henzel W, Goodman CS, Tessier-Lavigne M, Kidd T. Slit proteins bind Robo receptors and have an evolutionarily conserved role in repulsive axon guidance. Cell. 1999;96:795–806. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases