

Genetic and functional analyses of the conserved C-terminal core domain of Escherichia coli FtsZ
- PMID: 10601211
- PMCID: PMC94211
- DOI: 10.1128/JB.181.24.7531-7544.1999
Genetic and functional analyses of the conserved C-terminal core domain of Escherichia coli FtsZ
Abstract
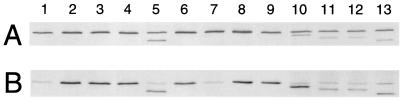
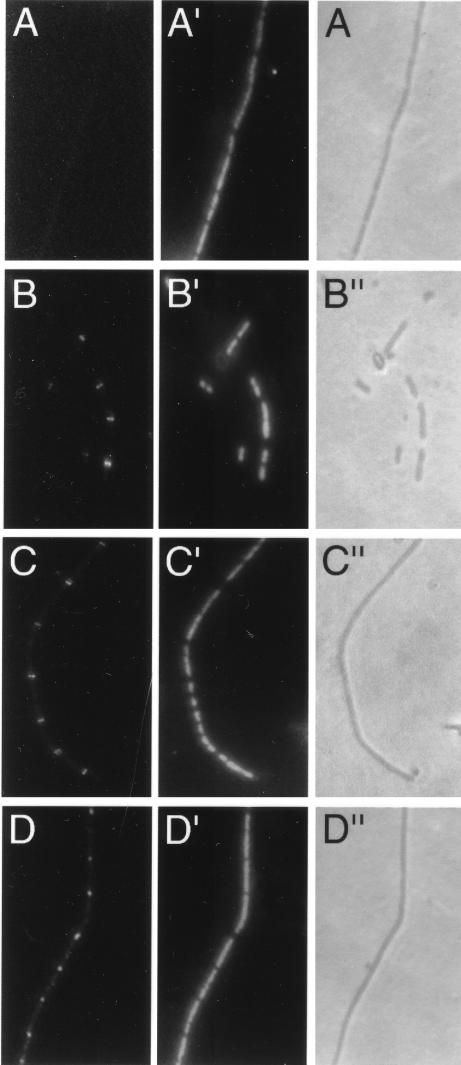
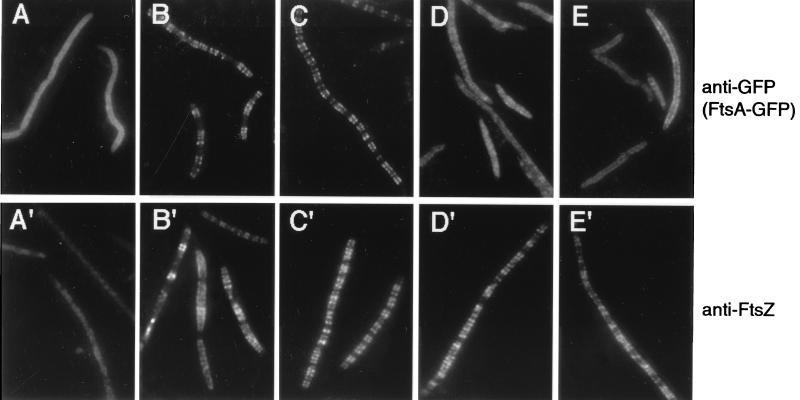
In Escherichia coli, FtsZ is required for the recruitment of the essential cell division proteins FtsA and ZipA to the septal ring. Several C-terminal deletions of E. coli FtsZ, including one of only 12 amino acids that removes the highly conserved C-terminal core domain, failed to complement chromosomal ftsZ mutants when expressed on a plasmid. To identify key individual residues within the core domain, six highly conserved residues were replaced with alanines. All but one of these mutants (D373A) failed to complement an ftsZ chromosomal mutant. Immunoblot analysis demonstrated that whereas I374A and F377A proteins were unstable in the cell, L372A, D373A, P375A, and L378A proteins were synthesized at normal levels, suggesting that they were specifically defective in some aspect of FtsZ function. In addition, all four of the stable mutant proteins were able to localize and form rings at potential division sites in chromosomal ftsZ mutants, implying a defect in a function other than localization and multimerization. Because another proposed function of FtsZ is the recruitment of FtsA and ZipA, we tested whether the C-terminal core domain was important for interactions with these proteins. Using two different in vivo assays, we found that the 12-amino-acid truncation of FtsZ was defective in binding to FtsA. Furthermore, two point mutants in this region (L372A and P375A) showed weakened binding to FtsA. In contrast, ZipA was capable of binding to all four stable point mutants in the FtsZ C-terminal core but not to the 12-amino-acid deletion.
Figures




References
-
- Addinall S G, Cao C, Lutkenhaus J. Temperature shift experiments with an ftsZ84(Ts) strain reveal rapid dynamics of FtsZ localization and indicate that the Z ring is required throughout septation and cannot reoccupy division sites once constriction has initiated. J Bacteriol. 1997;179:4277–4284. - PMC - PubMed
-
- Addinall S G, Lutkenhaus J. FtsZ-spirals and -arcs determine the shape of the invaginating septa in some mutants of Escherichia coli. Mol Microbiol. 1996;22:231–237. - PubMed
-
- Bai C, Elledge S J. Gene identification using the yeast two-hybrid system. Methods Enzymol. 1997;283:141–156. - PubMed
-
- Bartel P L, Fields S. Analyzing protein-protein interactions using two-hybrid system. Methods Enzymol. 1995;254:241–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
