

A nuclear 3'-5' exonuclease involved in mRNA degradation interacts with Poly(A) polymerase and the hnRNA protein Npl3p
- PMID: 10611239
- PMCID: PMC85144
- DOI: 10.1128/MCB.20.2.604-616.2000
A nuclear 3'-5' exonuclease involved in mRNA degradation interacts with Poly(A) polymerase and the hnRNA protein Npl3p
Abstract
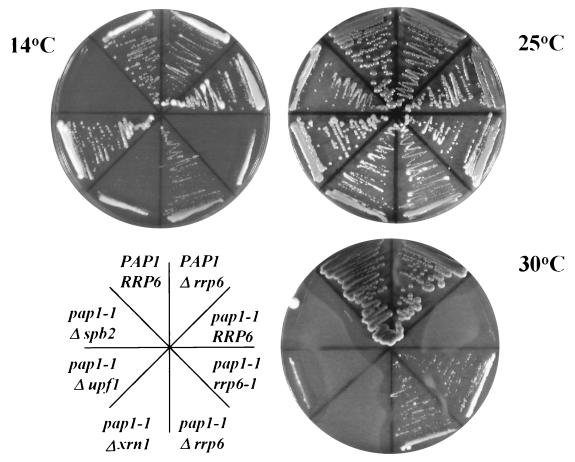
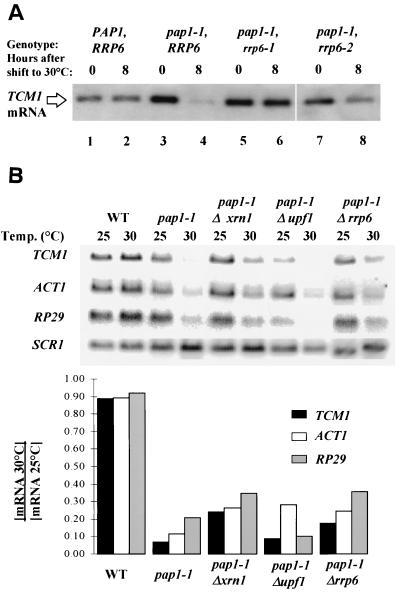
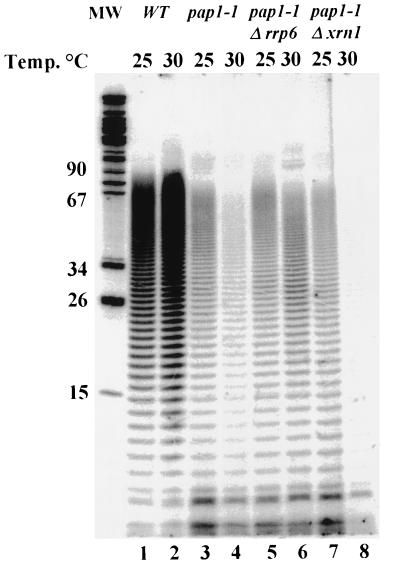
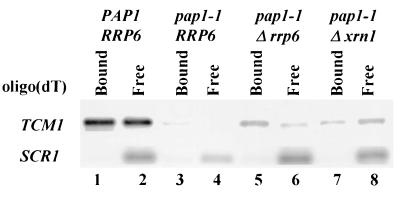
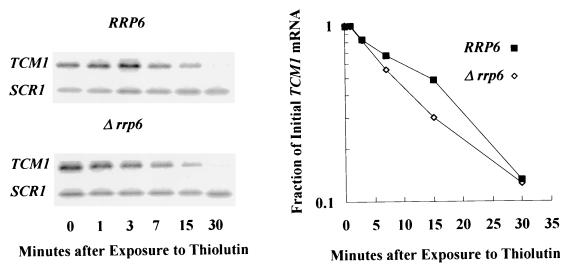
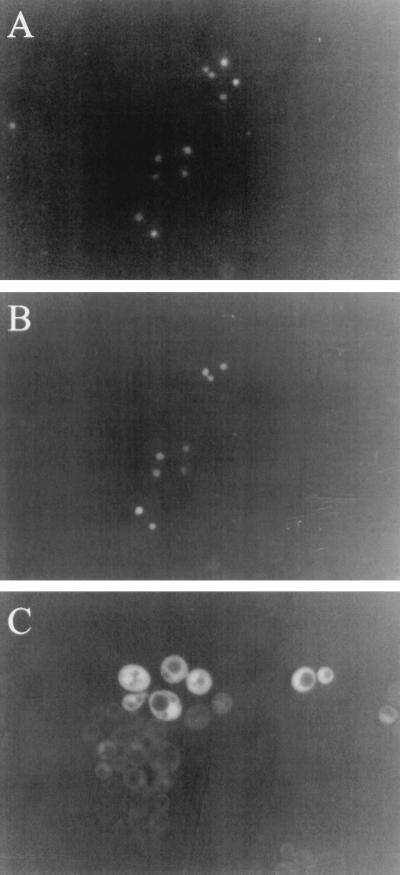
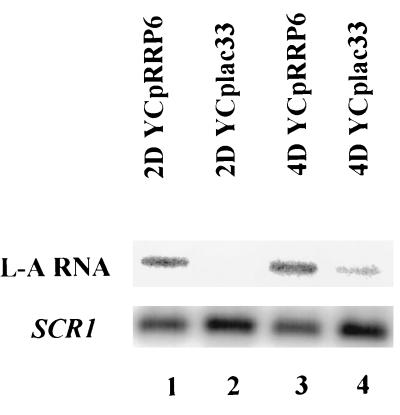
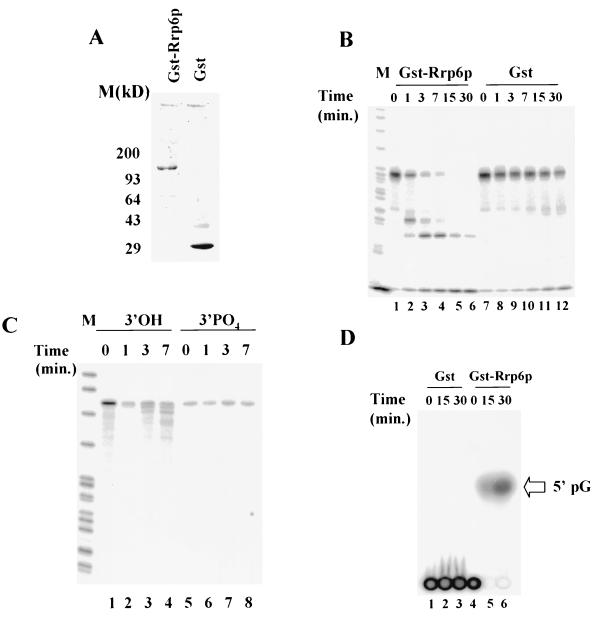
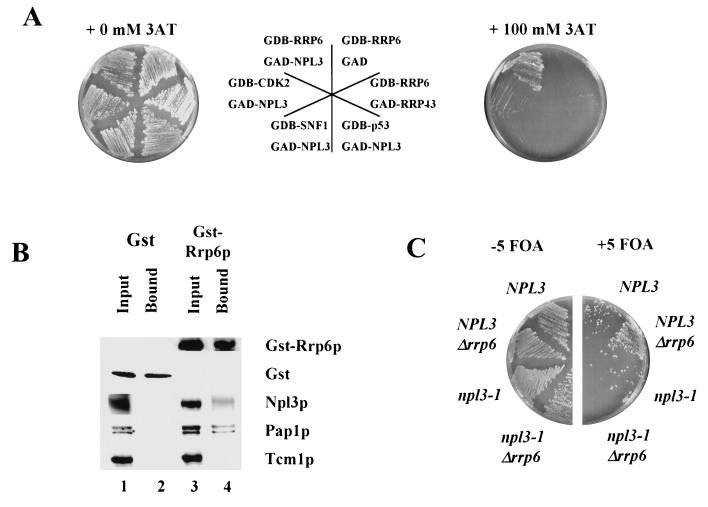
Inactivation of poly(A) polymerase (encoded by PAP1) in Saccharomyces cerevisiae cells carrying the temperature-sensitive, lethal pap1-1 mutation results in reduced levels of poly(A)(+) mRNAs. Genetic selection for suppressors of pap1-1 yielded two recessive, cold-sensitive alleles of the gene RRP6. These suppressors, rrp6-1 and rrp6-2, as well as a deletion of RRP6, allow growth of pap1-1 strains at high temperature and partially restore the levels of poly(A)(+) mRNA in a manner distinct from the cytoplasmic mRNA turnover pathway and without slowing a rate-limiting step in mRNA decay. Subcellular localization of an Rrp6p-green fluorescent protein fusion shows that the enzyme residues in the nucleus. Phylogenetic analysis and the nature of the rrp6-1 mutation suggest the existence of a highly conserved 3'-5' exonuclease core domain within Rrp6p. As predicted, recombinant Rrp6p catalyzes the hydrolysis of a synthetic radiolabeled RNA in a manner consistent with a 3'-5' exonucleolytic mechanism. Genetic and biochemical experiments indicate that Rrp6p interacts with poly(A) polymerase and with Npl3p, a poly(A)(+) mRNA binding protein implicated in pre-mRNA processing and mRNA nuclear export. These findings suggest that Rrp6p may interact with the mRNA polyadenylation system and thereby play a role in a nuclear pathway for the degradation of aberrantly processed precursor mRNAs.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials