

A bcr-3 isoform of RARalpha-PML potentiates the development of PML-RARalpha-driven acute promyelocytic leukemia
- PMID: 10611345
- PMCID: PMC24780
- DOI: 10.1073/pnas.96.26.15103
A bcr-3 isoform of RARalpha-PML potentiates the development of PML-RARalpha-driven acute promyelocytic leukemia
Abstract
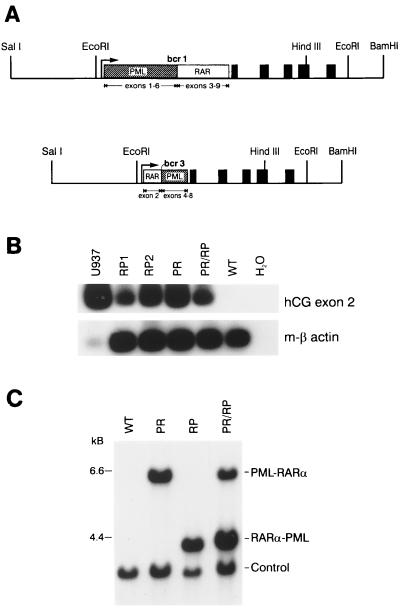
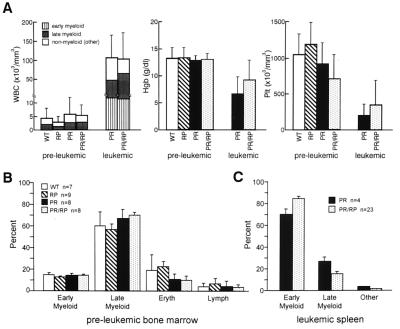
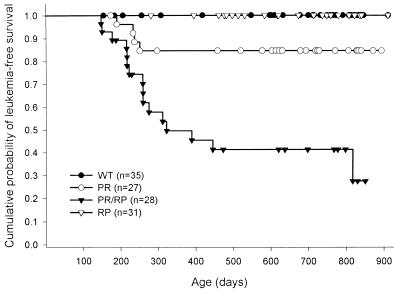
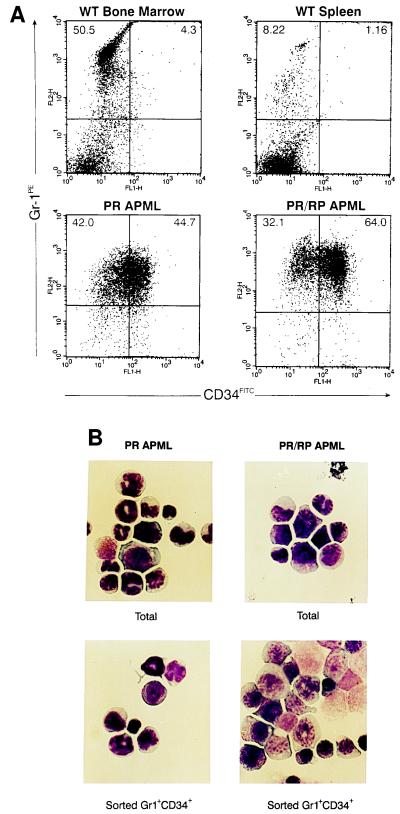
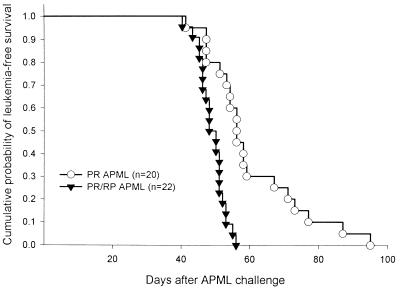
Acute promyelocytic leukemia (APML) most often is associated with the balanced reciprocal translocation t(15;17) (q22;q11.2) and the expression of both the PML-RARalpha and RARalpha-PML fusion cDNAs that are formed by this translocation. In this report, we investigated the biological role of a bcr-3 isoform of RARalpha-PML for the development of APML in a transgenic mouse model. Expression of RARalpha-PML alone in the early myeloid cells of transgenic mice did not alter myeloid development or cause APML, but its expression significantly increased the penetrance of APML in mice expressing a bcr-1 isoform of PML-RARalpha (15% of animals developed APML with PML-RARalpha alone vs. 57% with both transgenes, P < 0.001). The latency of APML development was not altered substantially by the expression of RARalpha-PML, suggesting that it does not behave as a classical "second hit" for development of the disease. Leukemias that arose from doubly transgenic mice were less mature than those from PML-RARalpha transgenic mice, but they both responded to all-trans retinoic acid in vitro. These findings suggest that PML-RARalpha drives the development of APML and defines its basic phenotype, whereas RARalpha-PML potentiates this phenotype via mechanisms that are not yet understood.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
