

Apolipoprotein E is essential for amyloid deposition in the APP(V717F) transgenic mouse model of Alzheimer's disease
- PMID: 10611368
- PMCID: PMC24803
- DOI: 10.1073/pnas.96.26.15233
Apolipoprotein E is essential for amyloid deposition in the APP(V717F) transgenic mouse model of Alzheimer's disease
Abstract
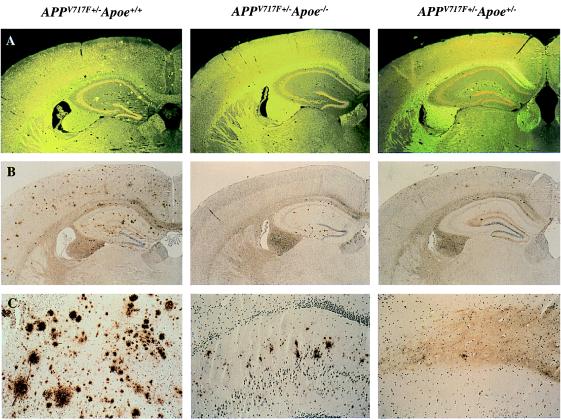
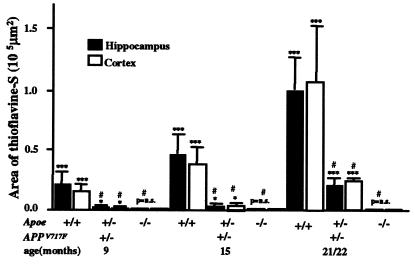
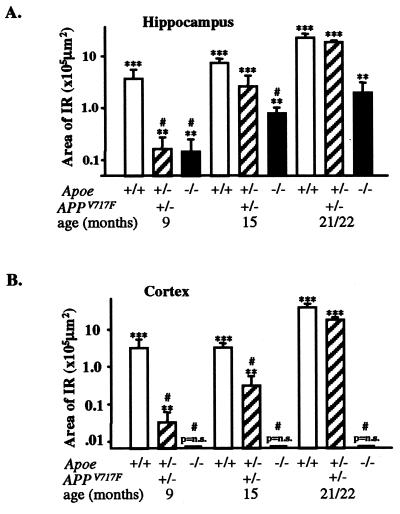
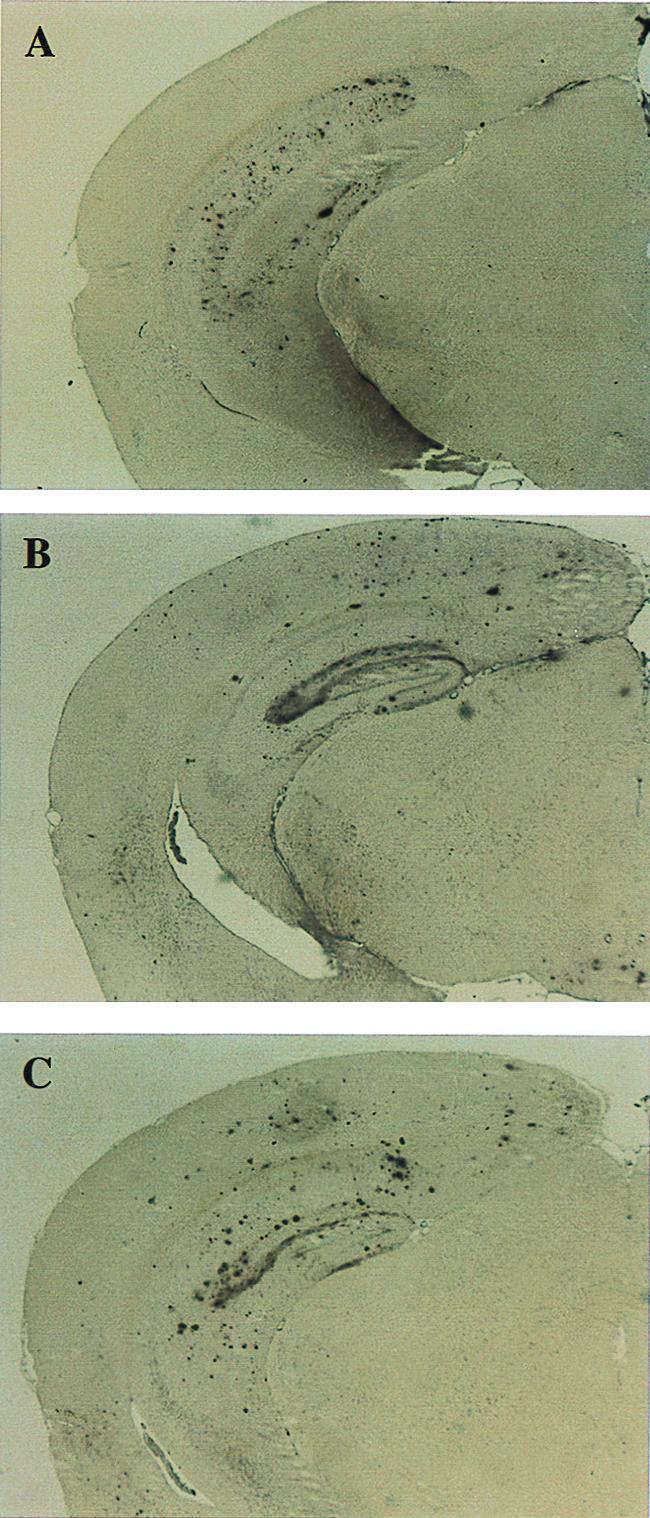
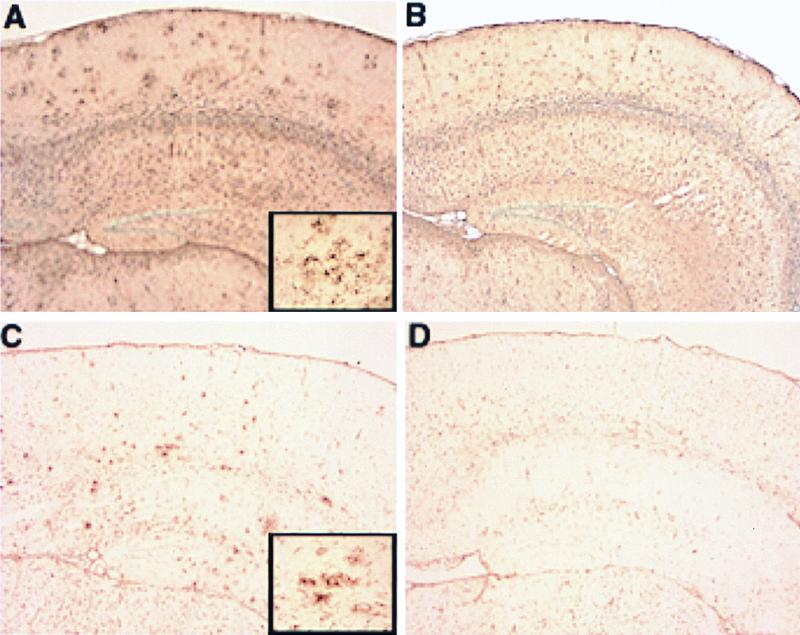
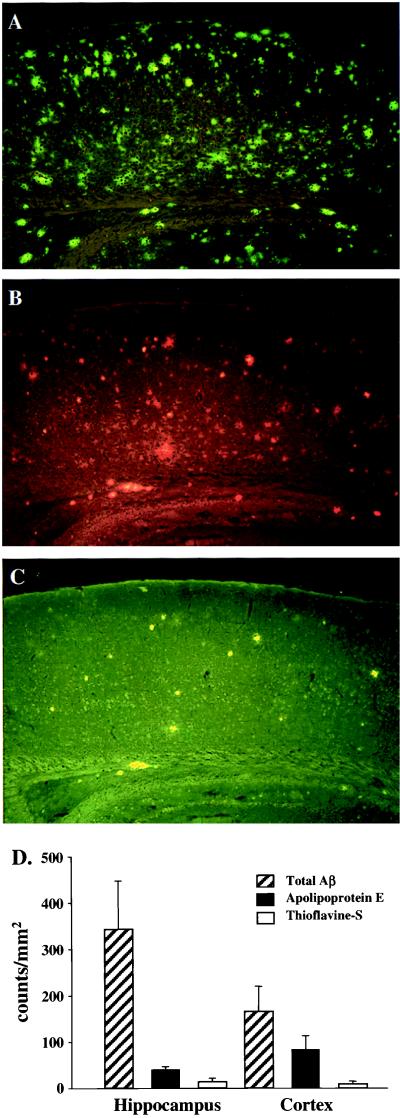
We quantified the amount of amyloid beta-peptide (Abeta) immunoreactivity as well as amyloid deposits in a large cohort of transgenic mice overexpressing the V717F human amyloid precursor protein (APP(V717F+/-) TG mice) with no, one, or two mouse apolipoprotein E (Apoe) alleles at various ages. Remarkably, no amyloid deposits were found in any brain region of APP(V717F+/-) Apoe(-/-) TG mice as old as 22 mo of age, whereas age-matched APP(V717F +/-) Apoe(+/-) and Apoe(+/+) TG mice display abundant amyloid deposition. The amount of Abeta immunoreactivity in the hippocampus was also markedly reduced in an Apoe gene dose-dependent manner (Apoe(+/+) > Apoe(+/-) >> Apoe(-/-)), and no Abeta immunoreactivity was detected in the cerebral cortex of APP(V717F+/-) Apoe(-/-) TG mice at any of the time points examined. The absence of apolipoprotein E protein (apoE) dramatically reduced the amount of both Abeta(1-40) and Abeta(1-42) immunoreactive deposits as well as the resulting astrogliosis and microgliosis normally observed in APP(V717F) TG mice. ApoE immunoreactivity was detected in a subset of Abeta immunoreactive deposits and in virtually all thioflavine-S-fluorescent amyloid deposits. Because the absence of apoE alters neither the transcription or translation of the APP(V717F) transgene nor its processing to Abeta peptide(s), we postulate that apoE promotes both the deposition and fibrillization of Abeta, ultimately affecting clearance of protease-resistant Abeta/apoE aggregates. ApoE appears to play an essential role in amyloid deposition in brain, one of the neuropathological hallmarks of Alzheimer's disease.
Figures
References
-
- Weisgraber K H, Mahley R W. FASEB J. 1996;10:1485–1494. - PubMed
-
- Corder E H, Saunders A M, Risch N J, Strittmatter W J, Schmechel D E, Gaskell P C, Jr, Rimmler J B, Locke P A, Conneally P M, Schmader K E, et al. Nat Genet. 1994;7:180–184. - PubMed
-
- Lambert J-C, Berr C, Pasquier F, Delacourte A, Frigard B, Cottel D, Pérez-Tur J, Mouroux V, Mohr M, Cécyre D, et al. Hum Mol Genet. 1998;7:1511–1516. - PubMed
-
- Bullido M J, Artiga M J, Recuero M, Sastre I, Garcia M A, Aldudo J, Lendon C, Han S W, Morris J C, Frank A, et al. Nat Genet. 1998;18:69–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
