

Abnormal neurotransmission in mice lacking synaptic vesicle protein 2A (SV2A)
- PMID: 10611374
- PMCID: PMC24809
- DOI: 10.1073/pnas.96.26.15268
Abnormal neurotransmission in mice lacking synaptic vesicle protein 2A (SV2A)
Abstract
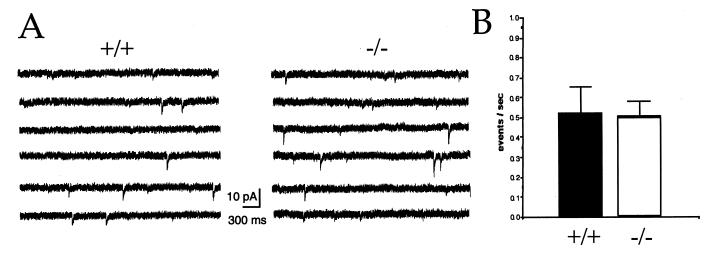
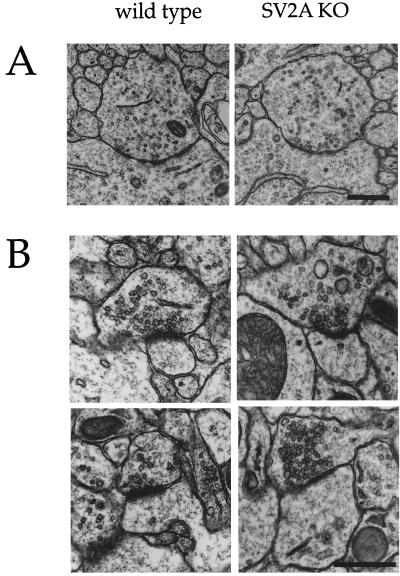
Synaptic vesicle protein 2 (SV2) is a membrane glycoprotein common to all synaptic and endocrine vesicles. Unlike many proteins involved in synaptic exocytosis, SV2 has no homolog in yeast, indicating that it performs a function unique to secretion in higher eukaryotes. Although the structure and protein interactions of SV2 suggest multiple possible functions, its role in synaptic events remains unknown. To explore the function of SV2 in an in vivo context, we generated mice that do not express the primary SV2 isoform, SV2A, by using targeted gene disruption. Animals homozygous for the SV2A gene disruption appear normal at birth. However, they fail to grow, experience severe seizures, and die within 3 weeks, suggesting multiple neural and endocrine deficits. Electrophysiological studies of spontaneous inhibitory neurotransmission in the CA3 region of the hippocampus revealed that loss of SV2A leads to a reduction in action potential-dependent gamma-aminobutyric acid (GABA)ergic neurotransmission. In contrast, action potential-independent neurotransmission was normal. Analyses of synapse ultrastructure suggest that altered neurotransmission is not caused by changes in synapse density or morphology. These findings demonstrate that SV2A is an essential protein and implicate it in the control of exocytosis.
Figures
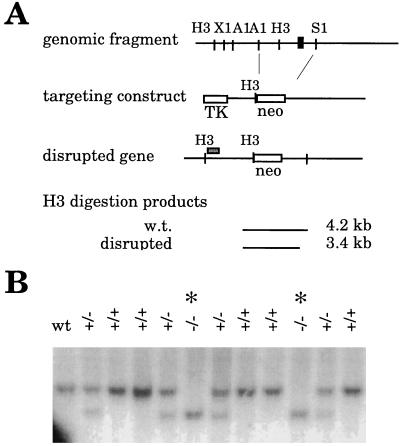
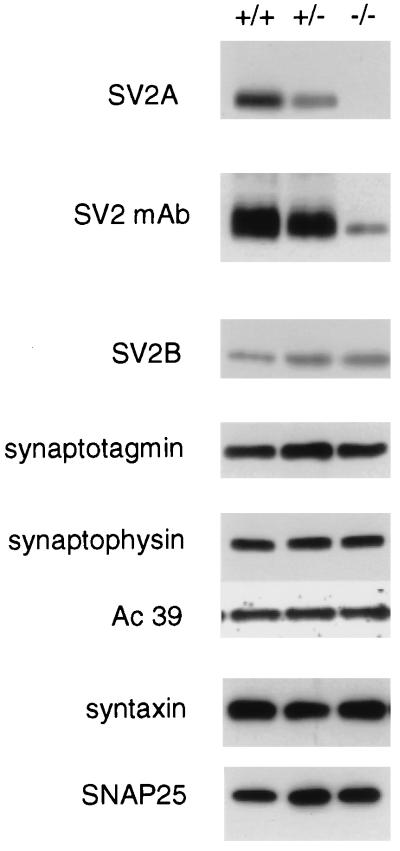
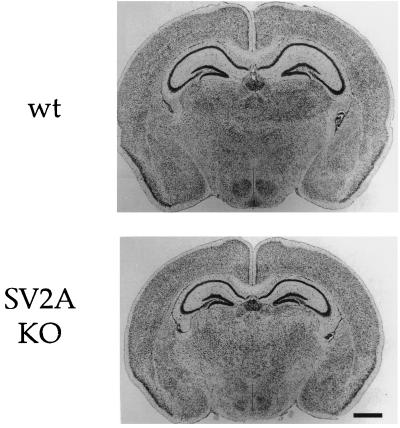
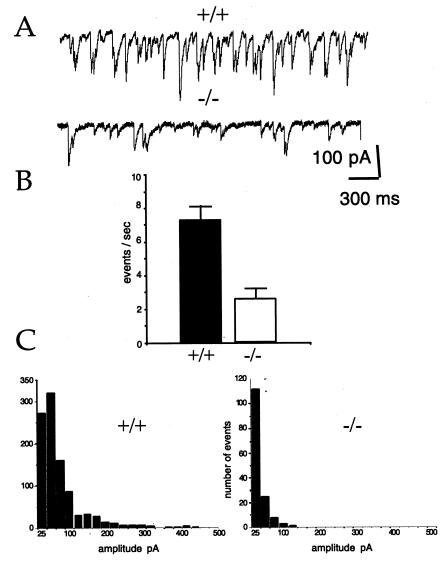
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
