

pH gating of ROMK (K(ir)1.1) channels: control by an Arg-Lys-Arg triad disrupted in antenatal Bartter syndrome
- PMID: 10611379
- PMCID: PMC24814
- DOI: 10.1073/pnas.96.26.15298
pH gating of ROMK (K(ir)1.1) channels: control by an Arg-Lys-Arg triad disrupted in antenatal Bartter syndrome
Abstract
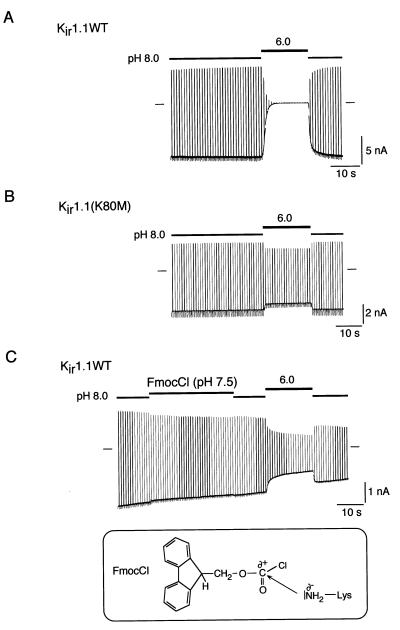
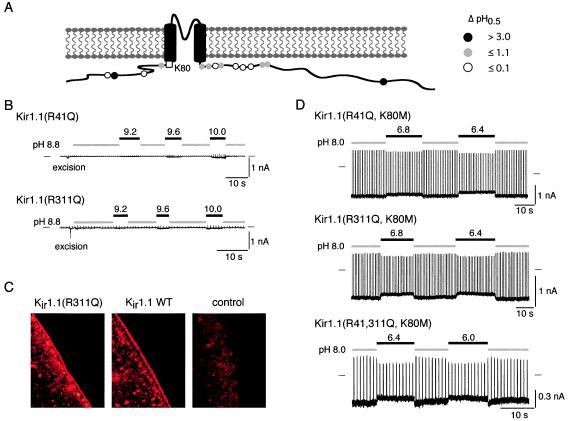
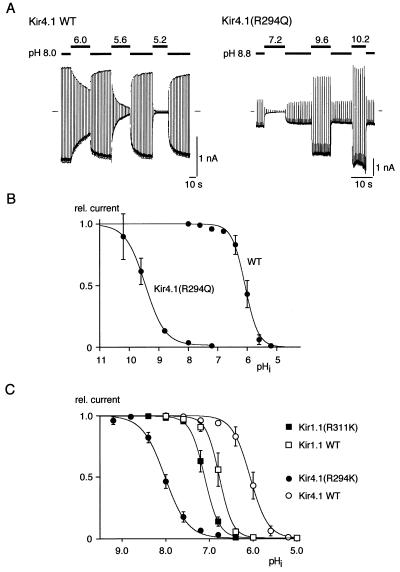
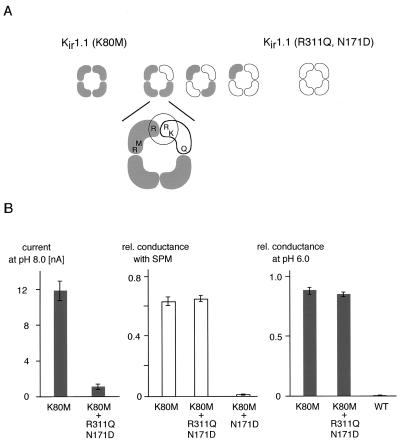
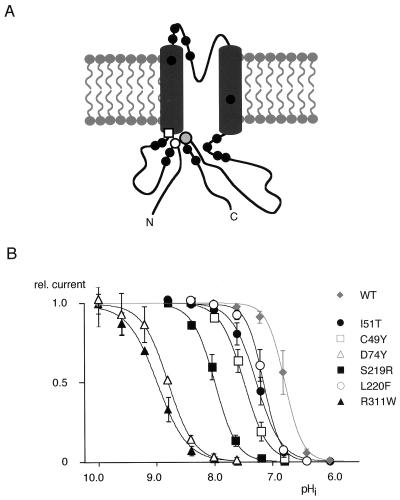
Inward-rectifier K(+) channels of the ROMK (K(ir)1.1) subtype are responsible for K(+) secretion and control of NaCl absorption in the kidney. A hallmark of these channels is their gating by intracellular pH in the neutral range. Here we show that a lysine residue close to TM1, identified previously as a structural element required for pH-induced gating, is protonated at neutral pH and that this protonation drives pH gating in ROMK and other K(ir) channels. Such anomalous titration of this lysine residue (Lys-80 in K(ir)1.1) is accomplished by the tertiary structure of the K(ir) protein: two arginines in the distant N and C termini of the same subunit (Arg-41 and Arg-311 in K(ir)1.1) are located in close spatial proximity to the lysine allowing for electrostatic interactions that shift its pK(a) into the neutral pH range. Structural disturbance of this triad as a result from a number of point mutations found in patients with antenatal Bartter syndrome shifts the pK(a) of the lysine residue off the neutral pH range and results in channels permanently inactivated under physiological conditions. Thus, the results provide molecular understanding for normal pH gating of K(ir) channels as well as for the channel defects found in patients with antenatal Bartter syndrome.
Figures
References
-
- Wang W, Sackin H, Giebisch G. Annu Rev Physiol. 1992;54:81–96. - PubMed
-
- Giebisch G. Am J Physiol. 1998;274:F817–F833. - PubMed
-
- Hebert S. Am J Physiol. 1998;275:F325–F327. - PubMed
-
- Ho K, Nichols C G, Lederer W J, Lytton J, Vassilev P M, Kanazirska M V, Hebert S C. Nature (London) 1993;362:31–38. - PubMed
-
- Nichols C, Lopatin A. Annu Rev Physiol. 1997;59:171–191. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
