

Intranuclear anchoring of repetitive DNA sequences: centromeres, telomeres, and ribosomal DNA
- PMID: 10613900
- PMCID: PMC2174248
- DOI: 10.1083/jcb.147.7.1409
Intranuclear anchoring of repetitive DNA sequences: centromeres, telomeres, and ribosomal DNA
Abstract
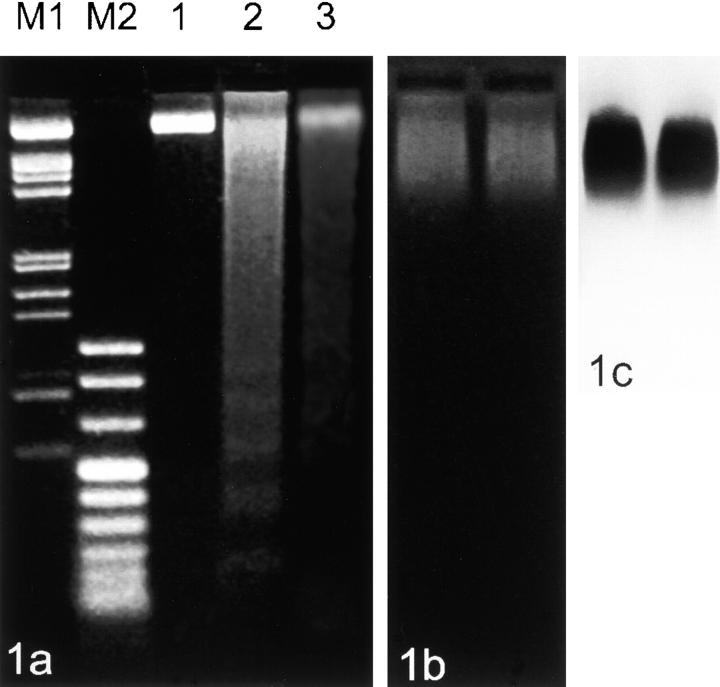
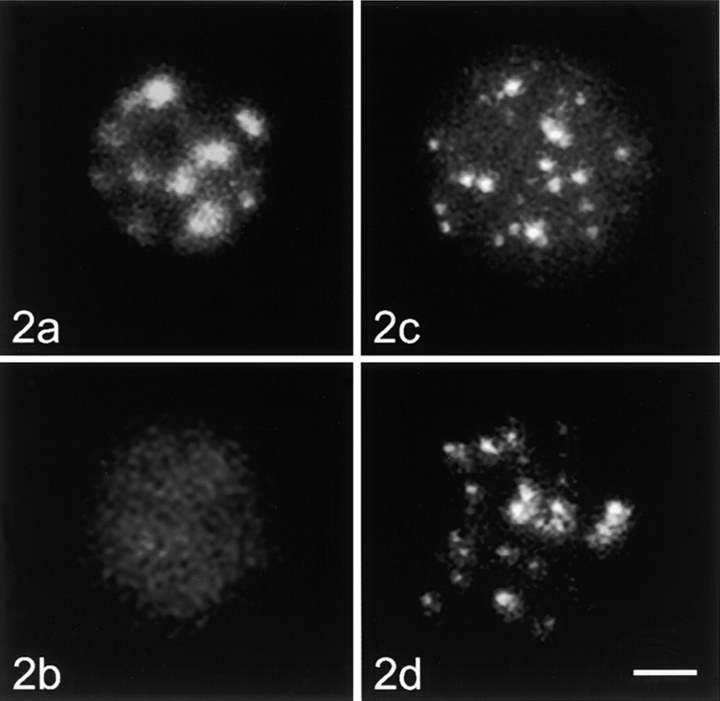
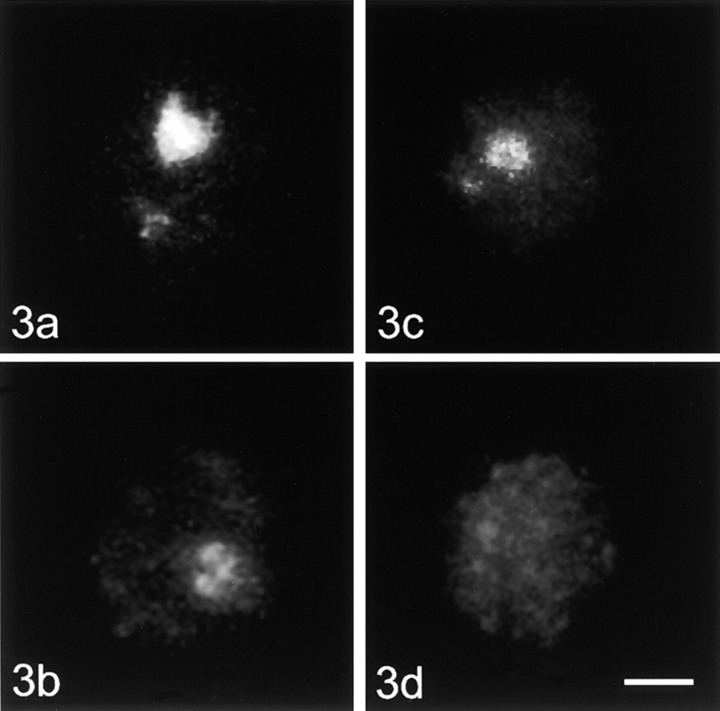
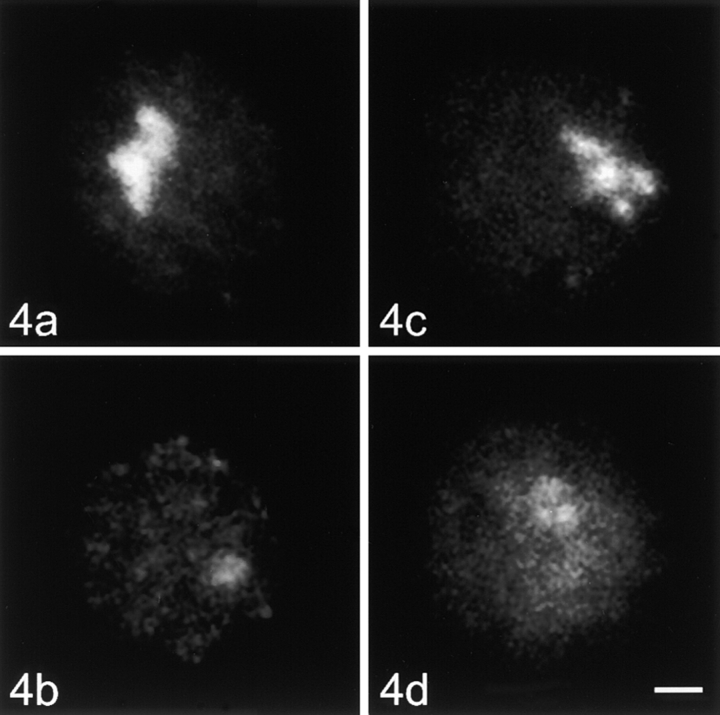
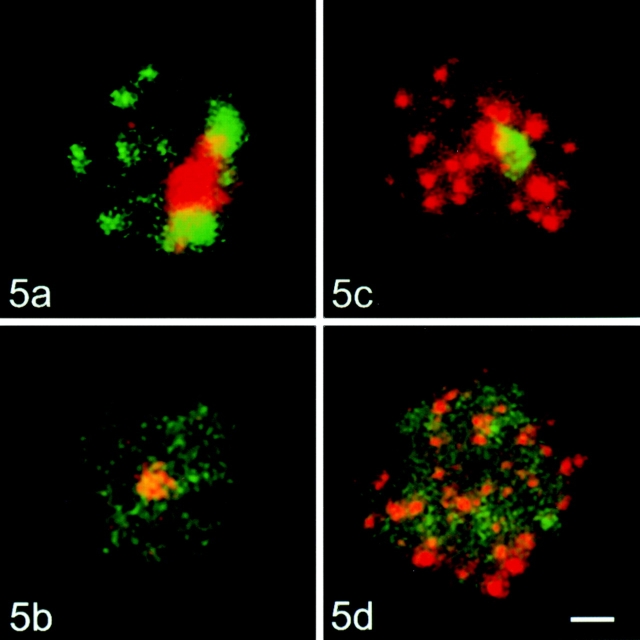
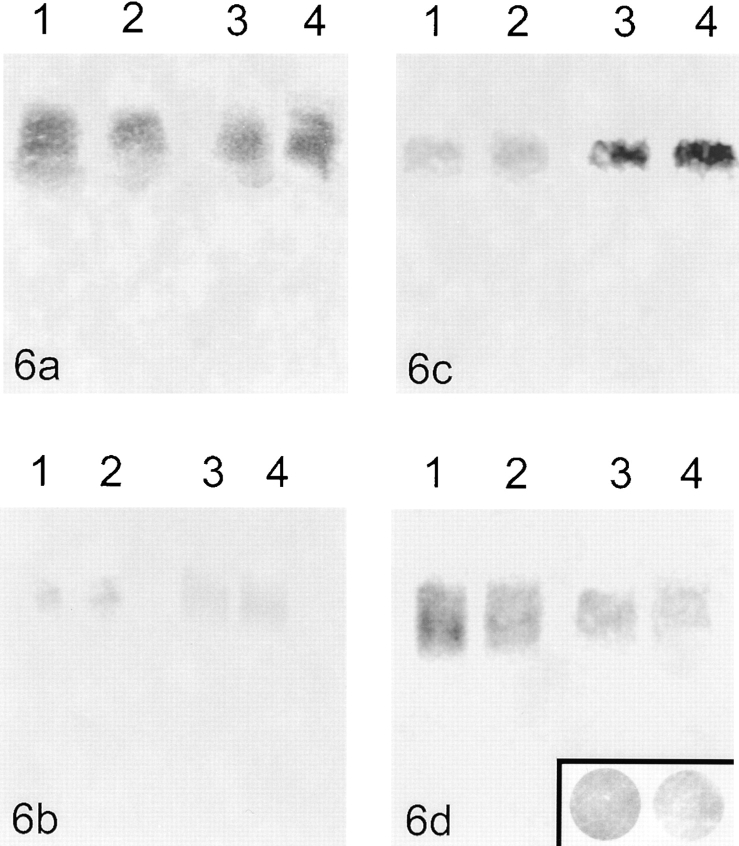
Centromeres, telomeres, and ribosomal gene clusters consist of repetitive DNA sequences. To assess their contributions to the spatial organization of the interphase genome, their interactions with the nucleoskeleton were examined in quiescent and activated human lymphocytes. The nucleoskeletons were prepared using "physiological" conditions. The resulting structures were probed for specific DNA sequences of centromeres, telomeres, and ribosomal genes by in situ hybridization; the electroeluted DNA fractions were examined by blot hybridization. In both nonstimulated and stimulated lymphocytes, centromeric alpha-satellite repeats were almost exclusively found in the eluted fraction, while telomeric sequences remained attached to the nucleoskeleton. Ribosomal genes showed a transcription-dependent attachment pattern: in unstimulated lymphocytes, transcriptionally inactive ribosomal genes located outside the nucleolus were eluted completely. When comparing transcription unit and intergenic spacer, significantly more of the intergenic spacer was removed. In activated lymphocytes, considerable but similar amounts of both rDNA fragments were eluted. The results demonstrate that: (a) the various repetitive DNA sequences differ significantly in their intranuclear anchoring, (b) telomeric rather than centromeric DNA sequences form stable attachments to the nucleoskeleton, and (c) different attachment mechanisms might be responsible for the interaction of ribosomal genes with the nucleoskeleton.
Figures
Similar articles
-
Cell cycle-dependent distribution of telomeres, centromeres, and chromosome-specific subsatellite domains in the interphase nucleus of mouse lymphocytes.Exp Cell Res. 1993 Mar;205(1):142-51. doi: 10.1006/excr.1993.1068. Exp Cell Res. 1993. PMID: 8453988
-
Arrangement of individual human ribosomal DNA fragments on stretched DNA fibers.Histochem Cell Biol. 1998 Aug;110(2):201-5. doi: 10.1007/s004180050282. Histochem Cell Biol. 1998. PMID: 9720993
-
Telomere and 45S rDNA sequences are structurally linked on the chromosomes in Chrysanthemum segetum L.Protoplasma. 2012 Jan;249(1):207-15. doi: 10.1007/s00709-011-0279-0. Epub 2011 May 3. Protoplasma. 2012. PMID: 21537919
-
Centromeres and telomeres.Curr Opin Cell Biol. 1992 Jun;4(3):379-84. doi: 10.1016/0955-0674(92)90002-t. Curr Opin Cell Biol. 1992. PMID: 1497910 Review.
-
Centromere identity from the DNA point of view.Chromosoma. 2014 Aug;123(4):313-25. doi: 10.1007/s00412-014-0462-0. Epub 2014 Apr 25. Chromosoma. 2014. PMID: 24763964 Free PMC article. Review.
Cited by
-
Ribosomal DNA and the Nucleolus as Keystones of Nuclear Architecture, Organization, and Function.Trends Genet. 2019 Oct;35(10):710-723. doi: 10.1016/j.tig.2019.07.011. Epub 2019 Aug 22. Trends Genet. 2019. PMID: 31447250 Free PMC article. Review.
-
The 4q subtelomere harboring the FSHD locus is specifically anchored with peripheral heterochromatin unlike most human telomeres.J Cell Biol. 2004 Oct 25;167(2):269-79. doi: 10.1083/jcb.200403128. J Cell Biol. 2004. PMID: 15504910 Free PMC article.
-
Dynamics of telomeres and promyelocytic leukemia nuclear bodies in a telomerase-negative human cell line.Mol Biol Cell. 2009 Apr;20(7):2070-82. doi: 10.1091/mbc.e08-02-0108. Epub 2009 Feb 11. Mol Biol Cell. 2009. PMID: 19211845 Free PMC article.
-
Clustering and protein dynamics of Drosophila melanogaster telomeres.Genetics. 2013 Oct;195(2):381-91. doi: 10.1534/genetics.113.155408. Epub 2013 Jul 26. Genetics. 2013. PMID: 23893488 Free PMC article.
-
Nuclear envelope tethering inhibits the formation of ALT-associated PML bodies in ALT cells.Aging (Albany NY). 2021 Apr 4;13(7):10490-10516. doi: 10.18632/aging.202810. Epub 2021 Apr 4. Aging (Albany NY). 2021. PMID: 33820871 Free PMC article.
References
-
- Allsopp R.C., Chang E., Kashefi-Aazam M., Rogaev E.I., Piatyszek M.A., Shay J.W., Harley C.B. Telomere shortening is associated with cell division in vitro and in vivo. Exp. Cell Res. 1995;220:194–200. - PubMed
-
- Allsopp R.C., Harley C.B. Evidence for a critical telomere length in senescent human fibroblasts. Exp. Cell Res. 1995;219:130–136. - PubMed
-
- Azzalin C.M., Mucciolo E., Bertoni L., Giulotto E. Fluorescence in situ hybridization with a synthetic (T2AG3)n polynucleotide detects several intrachromosomal telomere-like repeats on human chromosomes. Cytogenet. Cell Genet. 1997;78:112–115. - PubMed
-
- Barinaga M. Cells count proteins to keep their telomeres in line. Science. 1997;275:92. - PubMed
