

Developmental change in GABAA receptor desensitization kinetics and its role in synapse function in rat cortical neurons
- PMID: 10618148
- PMCID: PMC2269740
- DOI: 10.1111/j.1469-7793.2000.t01-5-00003.xm
Developmental change in GABAA receptor desensitization kinetics and its role in synapse function in rat cortical neurons
Abstract
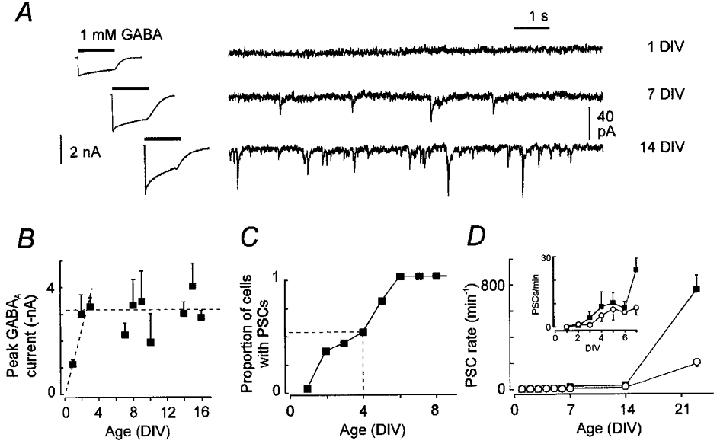
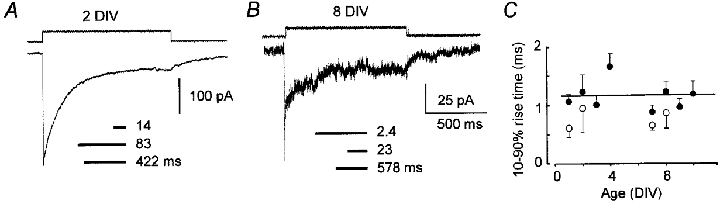
We examined the maturation of GABAA receptor synapses in cortical pyramidal neurons cultured from embryonic rats. The decay kinetics of GABAA receptor-mediated miniature postsynaptic currents (mPSCs) were compared with those of responses evoked by GABA in excised membrane patches. Fast perfusion of 1 or 10 mM GABA on membrane patches evoked currents with different desensitizing time courses in young and old neurons. For neurons older than 4 days in vitro (DIV), GABAA currents had a fast component of desensitization (median approximately 3 ms) seldom seen in patches from younger neurons. In contrast, mPSCs exhibited a substantial fast component of decay at 2-4 DIV that became more prominent with further development although the median value of its time constant remained unchanged. The selective alpha3 subunit positive modulator SB-205384 had no effect on mPSCs at any time in vitro but potentiated extrasynaptic activity. This suggests that synapse maturation does not proceed by a gradual exchange of early embryonic GABAA receptor subforms for adult forms. At all ages, the kinetic properties of mPSCs were heterogeneous. This heterogeneity extended to the level of mPSCs from single neurons and may be a normal aspect of synaptic functioning. These results suggest that inhibitory synapses in developing neurons are capable of selectively capturing GABAA receptors having fast desensitization kinetics. This functional preference probably reflects the developmental turning point from an inwardly looking trophic capacity of embryonic GABAA receptors to a role concerned with information processing.
Figures









Similar articles
-
Binge ethanol exposure delays development of GABAergic miniature postsynaptic currents in septal neurons.Brain Res Dev Brain Res. 2004 Sep 17;152(2):199-212. doi: 10.1016/j.devbrainres.2004.06.017. Brain Res Dev Brain Res. 2004. PMID: 15351508
-
GABA(A) receptor-mediated miniature postsynaptic currents and alpha-subunit expression in developing cortical neurons.J Neurophysiol. 1999 Dec;82(6):3286-97. doi: 10.1152/jn.1999.82.6.3286. J Neurophysiol. 1999. PMID: 10601460
-
Spontaneous GABAA receptor-mediated inhibitory currents in adult rat somatosensory cortex.J Neurophysiol. 1996 Apr;75(4):1573-88. doi: 10.1152/jn.1996.75.4.1573. J Neurophysiol. 1996. PMID: 8727397
-
Retrograde signaling at GABAA-receptor synapses in the mammalian CNS.Trends Neurosci. 1995 Aug;18(8):333-40. doi: 10.1016/0166-2236(95)93923-l. Trends Neurosci. 1995. PMID: 7482793 Review.
-
Short-term modulation of GABAA receptor function in the adult female rat.Prog Brain Res. 2002;139:31-42. doi: 10.1016/s0079-6123(02)39005-8. Prog Brain Res. 2002. PMID: 12436924 Review.
Cited by
-
Dynamic functions of GABA signaling during granule cell maturation.Front Neural Circuits. 2013 Jan 8;6:113. doi: 10.3389/fncir.2012.00113. eCollection 2012. Front Neural Circuits. 2013. PMID: 23316139 Free PMC article.
-
Fast IPSCs in rat thalamic reticular nucleus require the GABAA receptor beta1 subunit.J Physiol. 2006 Apr 15;572(Pt 2):459-75. doi: 10.1113/jphysiol.2006.106617. Epub 2006 Feb 9. J Physiol. 2006. PMID: 16469775 Free PMC article.
-
Developmental changes in P2X purinoceptors on glycinergic presynaptic nerve terminals projecting to rat substantia gelatinosa neurones.J Physiol. 2001 Oct 15;536(Pt 2):505-19. doi: 10.1111/j.1469-7793.2001.0505c.xd. J Physiol. 2001. PMID: 11600685 Free PMC article.
-
Region-specific developmental specialization of GABA-glycine cosynapses in laminas I-II of the rat spinal dorsal horn.J Neurosci. 2001 Oct 15;21(20):7871-80. doi: 10.1523/JNEUROSCI.21-20-07871.2001. J Neurosci. 2001. PMID: 11588160 Free PMC article.
-
α2-containing GABAA receptors expressed in hippocampal region CA3 control fast network oscillations.J Physiol. 2013 Feb 15;591(4):845-58. doi: 10.1113/jphysiol.2012.243725. Epub 2012 Oct 29. J Physiol. 2013. PMID: 23109109 Free PMC article.
References
-
- Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa JL. GABAA, NMDA and AMPA receptors: a developmentally regulated ‘menage a trois’. Trends in Neurosciences. 1997;20:523–529. - PubMed
-
- Brooks KA, Jin H, Price M, Dichter MA. Developmental expression of GABA(A) receptor subunit mRNAs in individual hippocampal neurons in vitro and in vivo. Journal of Neurochemistry. 1997;70:1017–1028. - PubMed
-
- Dekoninck Y, Mody I. Noise analysis of miniature IPSCs in adult rat brain slices: properties and modulation of synaptic GABAA receptor channels. Journal of Neurophysiology. 1994;71:1318–1335. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources