

Inactive conformation of the serpin alpha(1)-antichymotrypsin indicates two-stage insertion of the reactive loop: implications for inhibitory function and conformational disease
- PMID: 10618372
- PMCID: PMC26617
- DOI: 10.1073/pnas.97.1.67
Inactive conformation of the serpin alpha(1)-antichymotrypsin indicates two-stage insertion of the reactive loop: implications for inhibitory function and conformational disease
Abstract
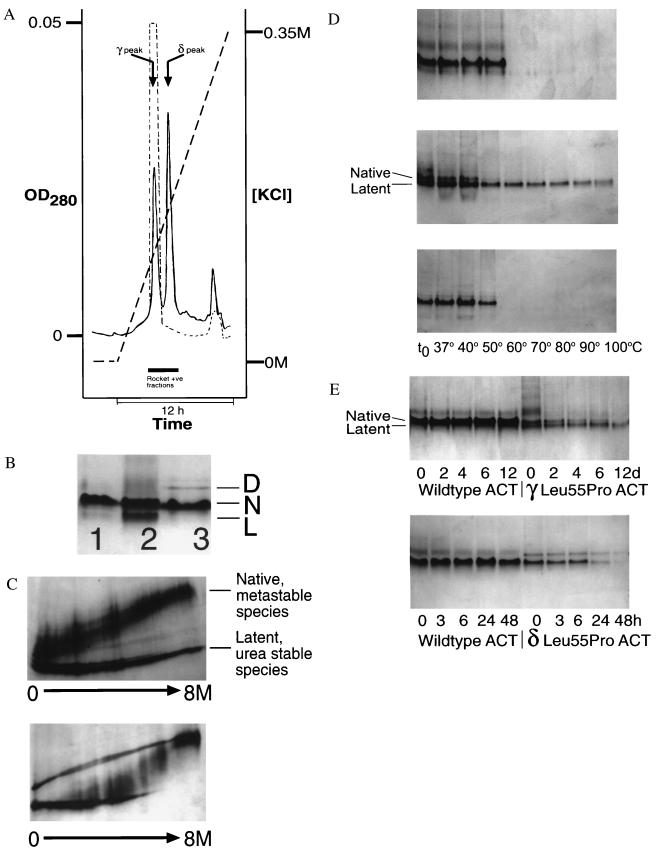
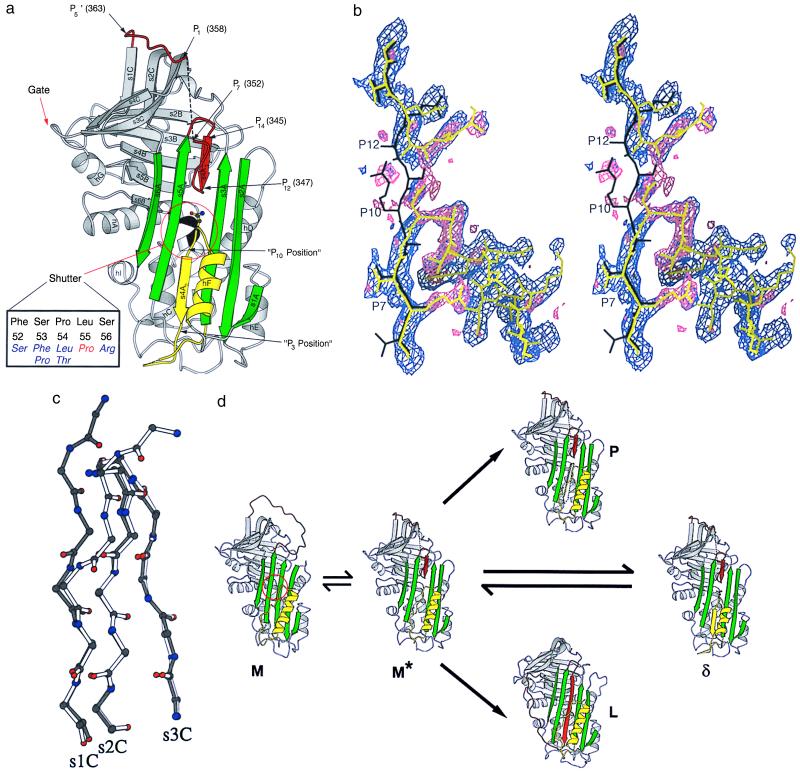
The serpins are a family of proteinase inhibitors that play a central role in the control of proteolytic cascades. Their inhibitory mechanism depends on the intramolecular insertion of the reactive loop into beta-sheet A after cleavage by the target proteinase. Point mutations within the protein can allow aberrant conformational transitions characterized by beta-strand exchange between the reactive loop of one molecule and beta-sheet A of another. These loop-sheet polymers result in diseases as varied as cirrhosis, emphysema, angio-oedema, and thrombosis, and we recently have shown that they underlie an early-onset dementia. We report here the biochemical characteristics and crystal structure of a naturally occurring variant (Leu-55-Pro) of the plasma serpin alpha(1)-antichymotrypsin trapped as an inactive intermediate. The structure demonstrates a serpin configuration with partial insertion of the reactive loop into beta-sheet A. The lower part of the sheet is filled by the last turn of F-helix and the loop that links it to s3A. This conformation matches that of proposed intermediates on the pathway to complex and polymer formation in the serpins. In particular, this intermediate, along with the latent and polymerized conformations, explains the loss of activity of plasma alpha(1)-antichymotrypsin associated with chronic obstructive pulmonary disease in patients with the Leu-55-Pro mutation.
Figures

References
-
- Aronsen K F, Ekelund G, Kindmark C O, Laurell C-B. Scand J Clin Lab Invest Suppl. 124. 1972;29:127–136. - PubMed
-
- Beatty K, Bieth J, Travis J. J Biol Chem. 1980;255:3931–3934. - PubMed
-
- Rubin H, Wang Z, Nickbarg E B, McLarney S, Naidoo N, Schoenberger O L, Johnson J L, Cooperman B S. J Biol Chem. 1990;265:1199–1207. - PubMed
-
- Lomas D A, Stone S R, Llewelyn-Jones C, Keogan M-T, Wang Z M, Rubin H, Carrell R W, Stockley R A. J Biol Chem. 1995;270:23437–23443. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
