

The crystal structure of modified bovine fibrinogen
- PMID: 10618375
- PMCID: PMC26620
- DOI: 10.1073/pnas.97.1.85
The crystal structure of modified bovine fibrinogen
Abstract
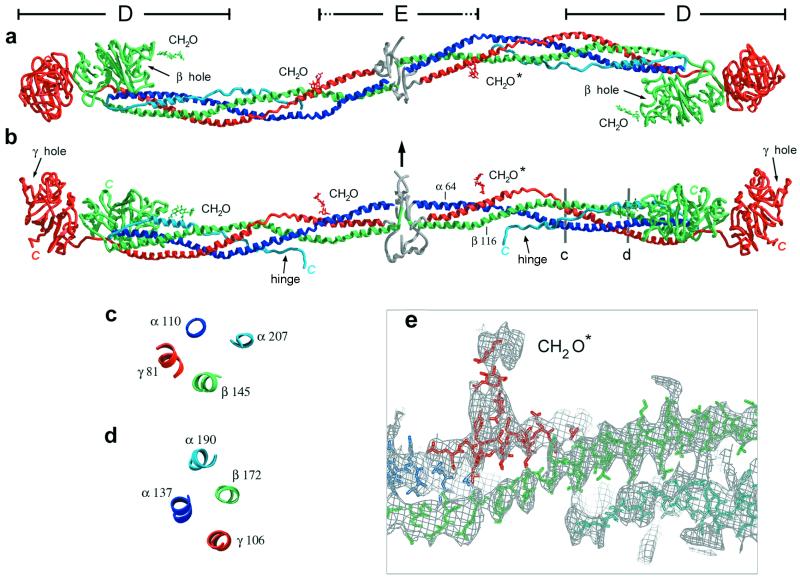
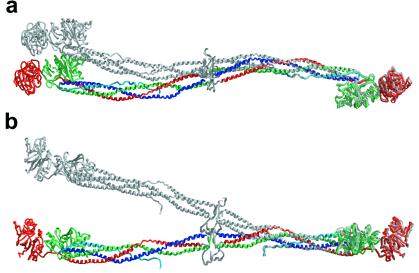
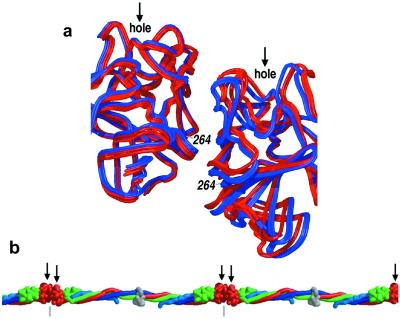
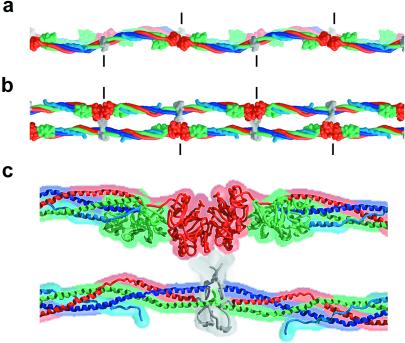
Here we report the crystal structure at approximately 4-A resolution of a selectively proteolyzed bovine fibrinogen. This key component in hemostasis is an elongated 340-kDa glycoprotein in the plasma that upon activation by thrombin self-assembles to form the fibrin clot. The crystals are unusual because they are made up of end-to-end bonded molecules that form flexible filaments. We have visualized the entire coiled-coil region of the molecule, which has a planar sigmoidal shape. The primary polymerization receptor pockets at the ends of the molecule face the same way throughout the end-to-end bonded filaments, and based on this conformation, we have developed an improved model of the two-stranded protofibril that is the basic building block in fibrin. Near the middle of the coiled-coil region, the plasmin-sensitive segment is a hinge about which the molecule adopts different conformations. This segment also includes the boundary between the three- and four-stranded portions of the coiled coil, indicating the location on the backbone that anchors the extended flexible Aalpha arm. We suggest that a flexible branch point in the molecule may help accommodate variability in the structure of the fibrin clot.
Figures
References
-
- Doolittle R F, Everse S J, Spraggon G. FASEB J. 1996;10:1464–1470. - PubMed
-
- Mosesson M W, Hainfeld J, Wall J, Haschemeyer R H. J Mol Biol. 1981;153:695–718. - PubMed
-
- Erickson H P, Fowler W E. In: Molecular Biology of Fibrinogen and Fibrin. Mosesson M W, Doolittle R F, editors. New York: N.Y. Acad. Sci.; 1983. pp. 146–163.
-
- Williams R C. In: Molecular Biology of Fibrinogen and Fibrin. Mosesson M W, Doolittle R F, editors. New York: N.Y. Acad. Sci.; 1983. pp. 180–193.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
