

Molecular cytogenetic dissection of human chromosomes 3 and 21 evolution
- PMID: 10618396
- PMCID: PMC26641
- DOI: 10.1073/pnas.97.1.206
Molecular cytogenetic dissection of human chromosomes 3 and 21 evolution
Abstract
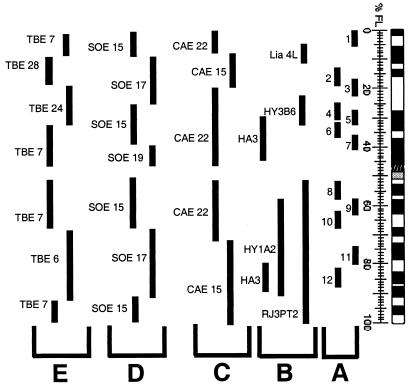
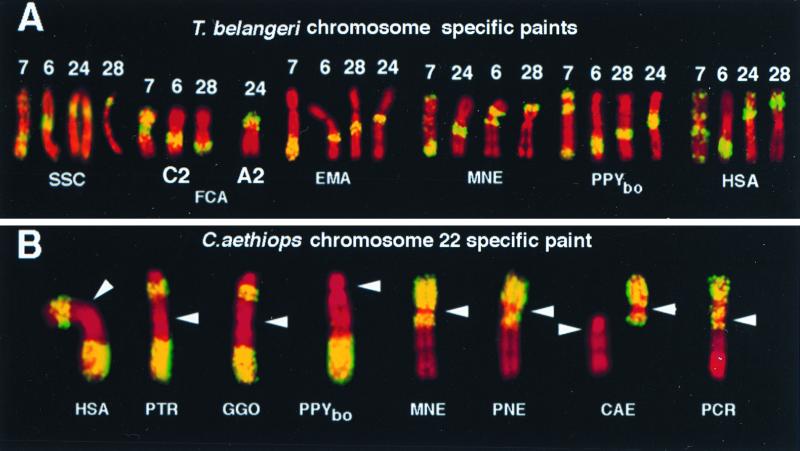
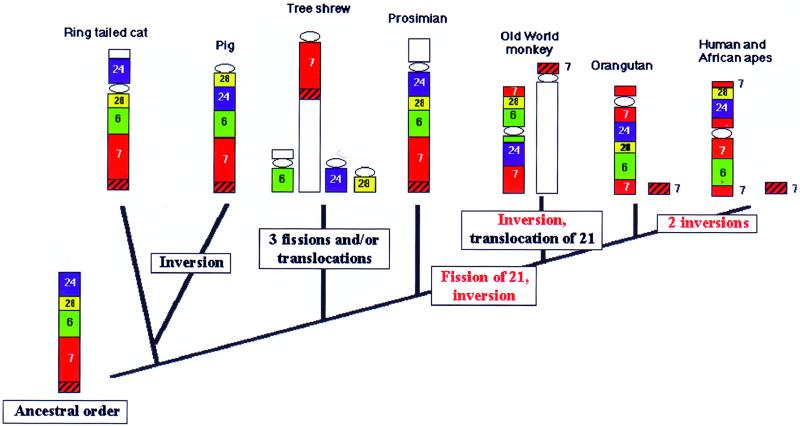
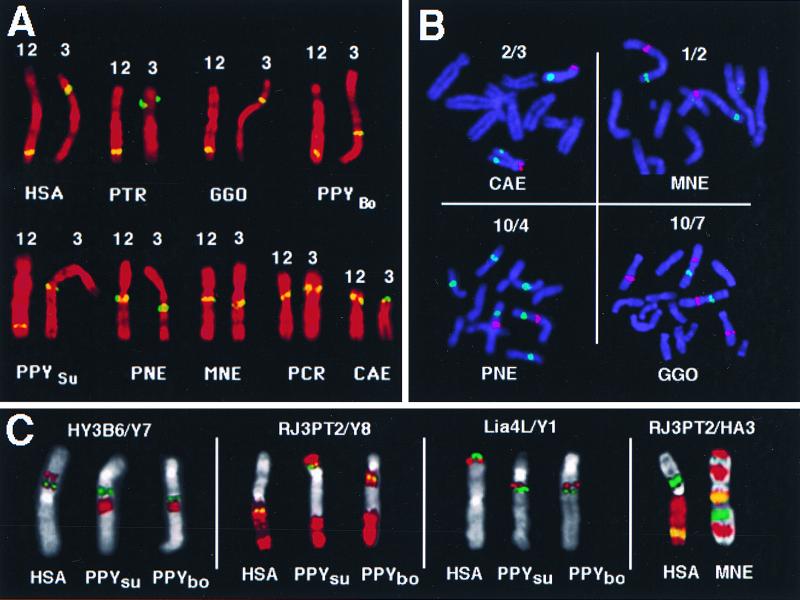
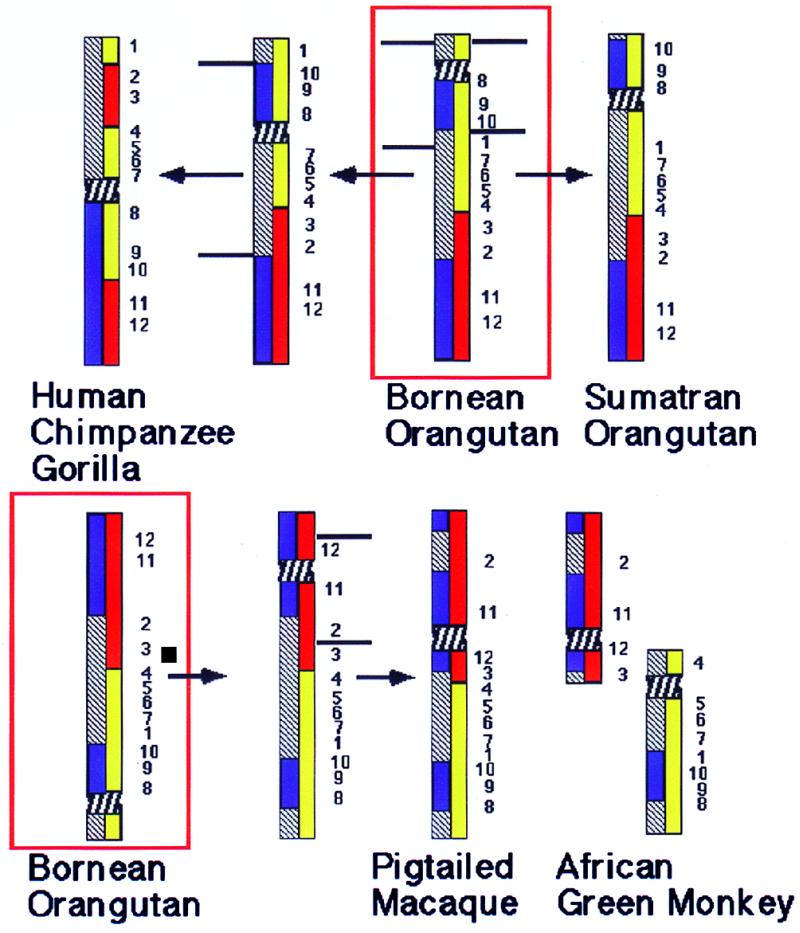
Chromosome painting in placental mammalians illustrates that genome evolution is marked by chromosomal synteny conservation and that the association of chromosomes 3 and 21 may be the largest widely conserved syntenic block known for mammals. We studied intrachromosomal rearrangements of the syntenic block 3/21 by using probes derived from chromosomal subregions with a resolution of up to 10-15 Mbp. We demonstrate that the rearrangements visualized by chromosome painting, mostly translocations, are only a fraction of the actual chromosomal changes that have occurred during evolution. The ancestral segment order for both primates and carnivores is still found in some species in both orders. From the ancestral primate/carnivore condition an inversion is needed to derive the pig homolog, and a fission of chromosome 21 and a pericentric inversion is needed to derive the Bornean orangutan condition. Two overlapping inversions in the chromosome 3 homolog then would lead to the chromosome form found in humans and African apes. This reconstruction of the origin of human chromosome 3 contrasts with the generally accepted scenario derived from chromosome banding in which it was proposed that only one pericentric inversion was needed. From the ancestral form for Old World primates (now found in the Bornean orangutan) a pericentric inversion and centromere shift leads to the chromosome ancestral for all Old World monkeys. Intrachromosomal rearrangements, as shown here, make up a set of potentially plentiful and informative markers that can be used for phylogenetic reconstruction and a more refined comparative mapping of the genome.
Figures
References
-
- O'Brien S J, Menotti-Raymond M, Murphy W J, Nash W G, Wienberg J, Stanyon R, Copland N G, Jenkins N A. Science. 1999;286:458–481. - PubMed
-
- Wienberg J, Stanyon R. Curr Opin Genet Dev. 1997;7:784–791. - PubMed
-
- Wienberg J, Stanyon R. Curr Opin Genet Dev. 1995;5:792–797. - PubMed
-
- Wienberg J, Stanyon R, Nash W G, O'Brien P C, Yang F, O'Brien S J, Ferguson-Smith M A. Cytogenet Cell Genet. 1997;77:211–217. - PubMed
-
- Richard F, Dutrillaux B. Chromosome Res. 1998;6:263–268. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
