

Differential roles of interleukin 15 mRNA isoforms generated by alternative splicing in immune responses in vivo
- PMID: 10620614
- PMCID: PMC2195806
- DOI: 10.1084/jem.191.1.157
Differential roles of interleukin 15 mRNA isoforms generated by alternative splicing in immune responses in vivo
Abstract
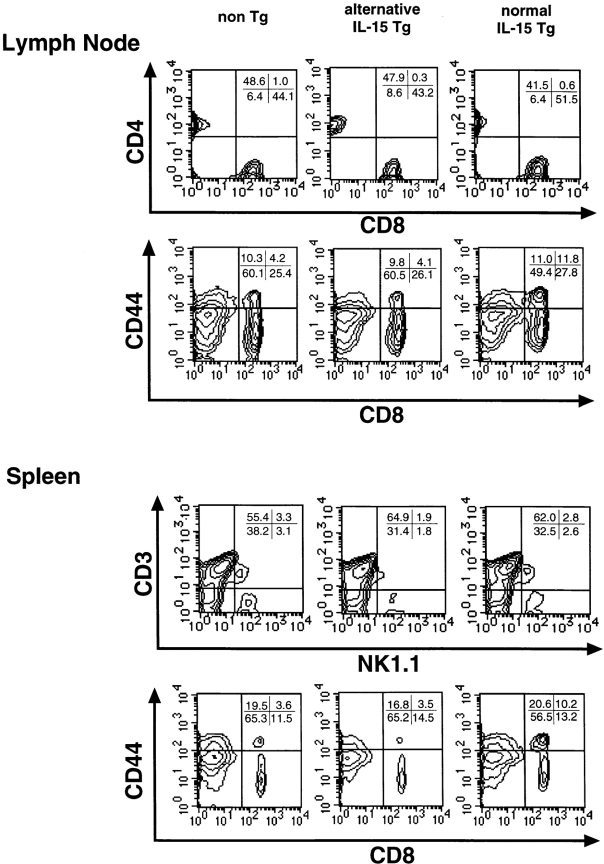
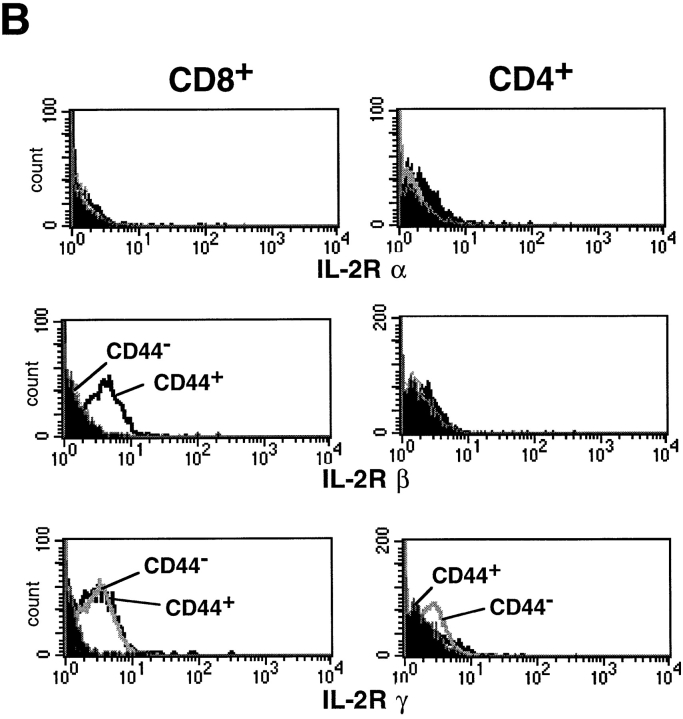
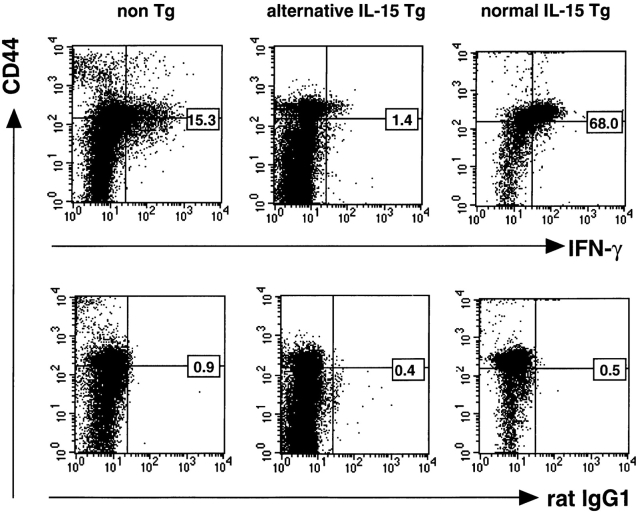
At least two types of interleukin (IL)-15 mRNA isoforms are generated by alternative splicing at the 5' upstream of exon 5 in mice. To elucidate the potential roles of IL-15 isoforms in immune responses in vivo, we constructed two groups of transgenic mice using originally described IL-15 cDNA with a normal exon 5 (normal IL-15 transgenic [Tg] mice) and IL-15 cDNA with an alternative exon 5 (alternative IL-15 Tg mice) under the control of an MHC class I promoter. Normal IL-15 Tg mice constitutionally produced a significant level of IL-15 protein and had markedly increased numbers of memory type (CD44(high) Ly6C(+)) of CD8(+) T cells in the LN. These mice showed resistance to Salmonella infection accompanied by the enhanced interferon (IFN)-gamma production, but depletion of CD8(+) T cells exaggerated the bacterial growth, suggesting that the IL-15-dependent CD8(+) T cells with a memory phenotype may serve to protect against Salmonella infection in normal IL-15 Tg mice. On the other hand, a large amount of intracellular IL-15 protein was detected but hardly secreted extracellularly in alternative IL-15 Tg mice. Although most of the T cells developed normally in the alternative IL-15 Tg mice, they showed impaired IFN-gamma production upon TCR engagement. The alternative IL-15 transgenic mice were susceptible to Salmonella accompanied by impaired production of endogenous IL-15 and IFN-gamma. Thus, two groups of IL-15 Tg mice may provide information concerning the different roles of IL-15 isoforms in the immune system in vivo.
Figures













References
-
- Tagaya Y., Bamford R.N., DeFilippis A., Waldmann T.A. IL-15a pleiotropic cytokine with diverse receptor/signaling pathways whose expression is controlled at multiple levels. Immunity. 1996;4:329–336 . - PubMed
-
- Waldmann T.A., Tagaya Y., Bamford R.N. Interleukin-2, interleukin-15, and their receptors. Int. Rev. Immunol. 1998;16:205–226 . - PubMed
-
- Waldmann T.A., Tagaya Y. The multifaceted regulation of interleukin-15 expression and the role of this cytokine in NK cell differentiation and host response to intracellular pathogens. Annu. Rev. Immunol. 1999;17:19–49 . - PubMed
-
- Grabstein K.H., Eisenmann J., Shanebeck K., Rauch C., Srinivasan S., Fung V., Beers C., Richardson J., Schoenborn M.A., Ahdieh M. Cloning of a T-cell growth factor that interacts with the β chain of the interleukin-2 receptor. Science. 1994;246:965–968 . - PubMed
-
- Bamford R.N., Battiata A.P., Burton J.D., Sharma H., Waldmann T.A. IL-15/T production by the adult T-cell leukemia cell line HuT-102 is associated with an HTLV-I-R/IL-15 fusion message that lacks many upstream AUGs that normally attenuate IL-15 mRNA translation. Proc. Natl. Acad. Sci. USA. 1996;93:2897–2902 . - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
