

Entry versus blockade of brain infection following oral or intraperitoneal scrapie administration: role of prion protein expression in peripheral nerves and spleen
- PMID: 10623745
- PMCID: PMC111603
- DOI: 10.1128/jvi.74.2.828-833.2000
Entry versus blockade of brain infection following oral or intraperitoneal scrapie administration: role of prion protein expression in peripheral nerves and spleen
Abstract
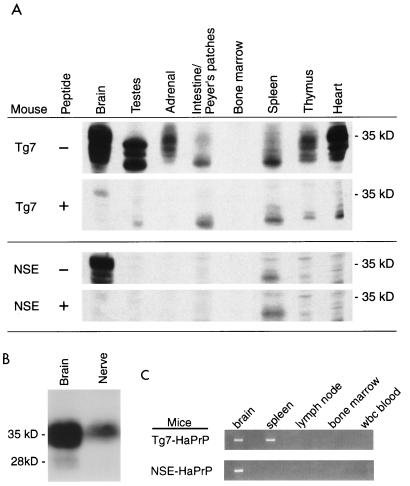
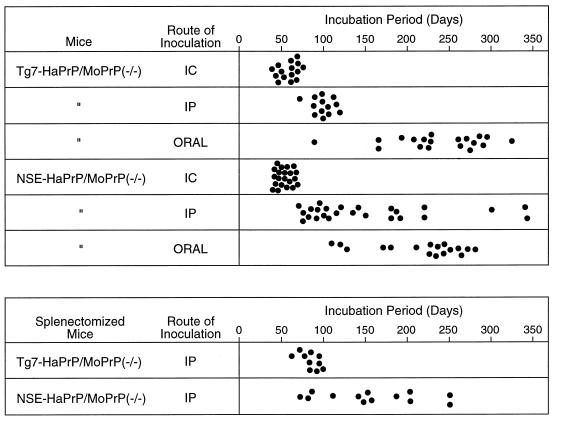
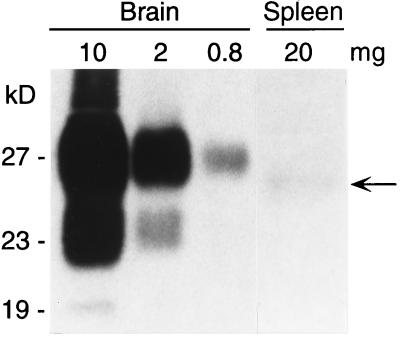
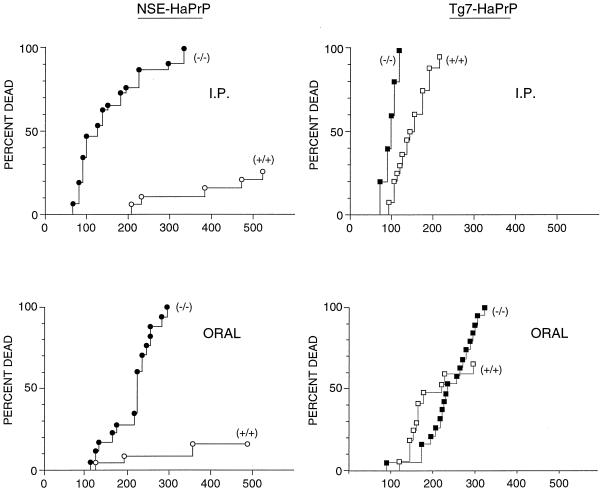
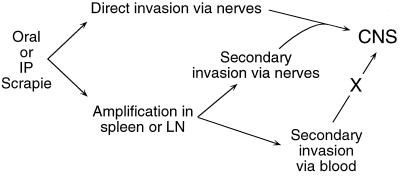
Naturally occurring transmissible spongiform encephalopathy (TSE) diseases such as bovine spongiform encephalopathy in cattle are probably transmitted by oral or other peripheral routes of infection. While prion protein (PrP) is required for susceptibility, the mechanism of spread of infection to the brain is not clear. Two prominent possibilities include hematogenous spread by leukocytes and neural spread by axonal transport. In the present experiments, following oral or intraperitoneal infection of transgenic mice with hamster scrapie strain 263K, hamster PrP expression in peripheral nerves was sufficient for successful infection of the brain, and cells of the spleen were not required either as a site of amplification or as transporters of infectivity. The role of tissue-specific PrP expression of foreign PrP in interference with scrapie infection was also studied in these transgenic mice. Peripheral expression of heterologous PrP completely protected the majority of mice from clinical disease after oral or intraperitoneal scrapie infection. Such extensive protection has not been seen in earlier studies on interference, and these results suggested that gene therapy with mutant PrP may be effective in preventing TSE diseases.
Figures
References
-
- Beekes M, McBride P A, Baldauf E. Cerebral targeting indicates vagal spread of infection in hamsters fed with scrapie. J Gen Virol. 1998;79:601–607. - PubMed
-
- Bendheim P E, Brown H R, Rudelli R D, Scala L J, Goller N L, Wen G Y, Kascsak R J, Cashman N R, Bolton D C. Nearly ubiquitous tissue distribution of the scrapie agent precursor protein. Neurology. 1992;42:149–156. - PubMed
-
- Blättler T, Brandner S, Raeber A J, Klein M A, Voigtlander T, Weissmann C, Aguzzi A. PrP-expressing tissue required for transfer of scrapie infectivity from spleen to brain. Nature. 1997;389:69–73. - PubMed
-
- Brandner S, Isenmann S, Raeber A, Fischer M, Sailer A, Kobayashi Y, Marino S, Weissmann C, Aguzzi A. Normal host prion protein necessary for scrapie-induced neurotoxicity. Nature. 1996;379:339–343. - PubMed
-
- Brown K L, Stewart K, Bruce M E, Fraser H. Severely combined immunodeficient (SCID) mice resist infection with bovine spongiform encephalopathy. J Gen Virol. 1997;78:2707–2710. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
