

Toward a more accurate quantitation of the activity of recombinant retroviruses: alternatives to titer and multiplicity of infection
- PMID: 10627536
- PMCID: PMC111460
- DOI: 10.1128/jvi.74.3.1258-1266.2000
Toward a more accurate quantitation of the activity of recombinant retroviruses: alternatives to titer and multiplicity of infection
Erratum in
-
Toward a more accurate quantitation of the activity of recombinant retroviruses: alternatives to titer and multiplicity of infection.J Virol. 2000 Apr;74(7):3431-9. doi: 10.1128/jvi.74.7.3431-3431.2000. J Virol. 2000. PMID: 10755888 Free PMC article.
Abstract
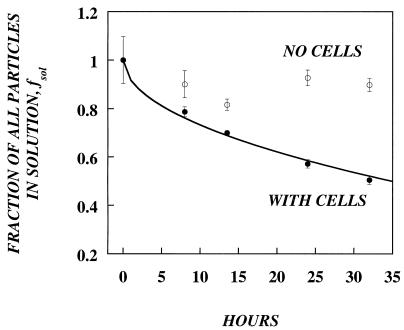
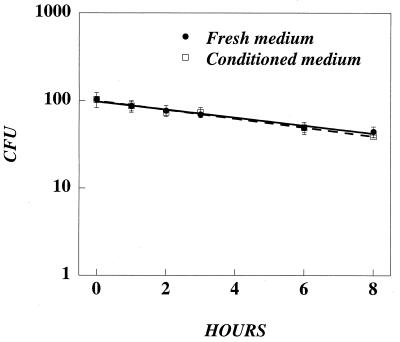
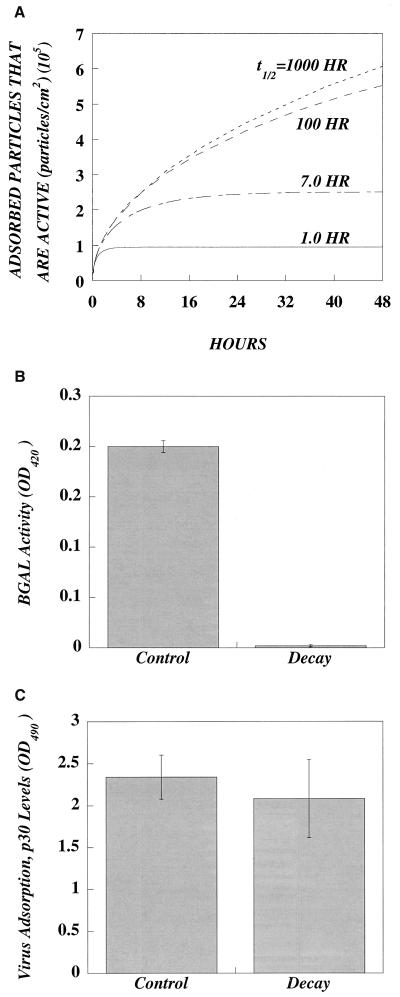
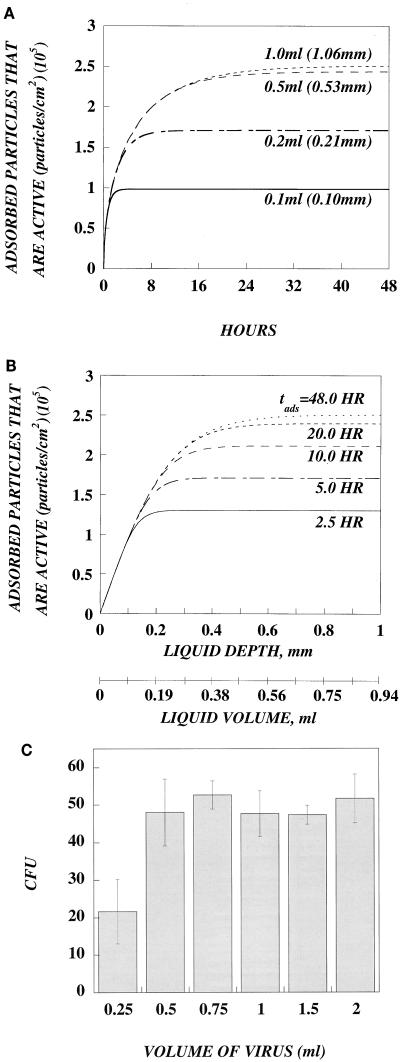
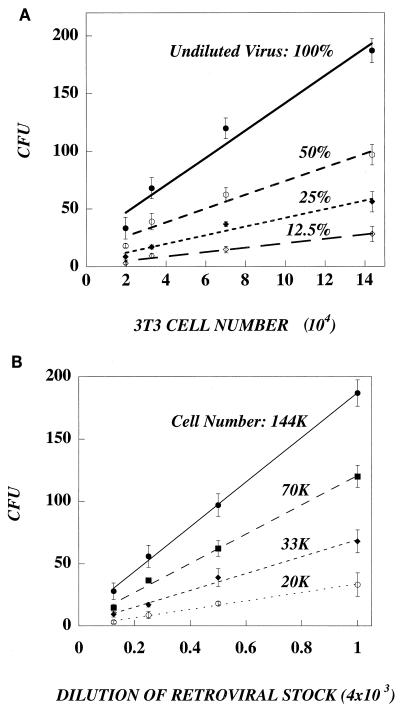
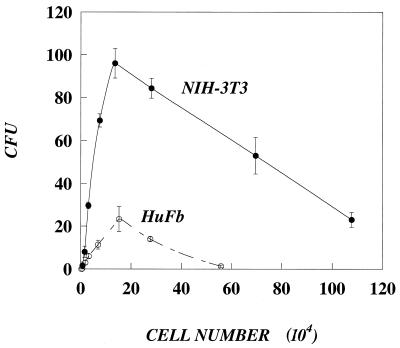
In this paper, we present a mathematical model with experimental support of how several key parameters govern the adsorption of active retrovirus particles onto the surface of adherent cells. These parameters, including time of adsorption, volume of virus, and the number, size, and type of target cells, as well as the intrinsic properties of the virus, diffusion coefficient, and half-life (t(1/2)), have been incorporated into a mathematical expression that describes the rate at which active virus particles adsorb to the cell surface. From this expression, we have obtained estimates of C(vo), the starting concentration of active retrovirus particles. In contrast to titer, C(vo) is independent of the specific conditions of the assay. The relatively slow diffusion (D = 2 x 10(-8) cm(2)/s) and rapid decay (t(1/2) = 6 to 7 h) of retrovirus particles explain why C(vo) values are significantly higher than titer values. Values of C(vo) also indicate that the number of defective particles in a retrovirus stock is much lower than previously thought, which has implications especially for the use of retroviruses for in vivo gene therapy. With this expression, we have also computed AVC (active viruses/cell), the number of active retrovirus particles that would adsorb per cell during a given adsorption time. In contrast to multiplicity of infection, which is based on titer and is subject to the same inaccuracies, AVC is based on the physicochemical parameters of the transduction assay and so is a more reliable alternative.
Figures
References
-
- Andreadis S, Fuller A O, Palsson B O. Cell cycle dependence of retroviral transduction: an issue of overlapping time scales. Biotechnol Bioeng. 1998;58:272–281. - PubMed
-
- Andreadis S, Palsson B O. Kinetics of retrovirus mediated gene transfer: the importance of the intracellular half-life of retroviruses. J Theor Biol. 1996;182:1–20. - PubMed
-
- Bahnson A B, Dunigan J T, Baysal B E, Mohney T, Atchison R W, Nimgaonkar M T, Ball E D, Barranger J A. Centrifugal enhancement of retroviral mediated gene transfer. J Virol Methods. 1995;54:131–143. - PubMed
-
- Chuck A S, Clarke M F, Palsson B O. Retroviral infection is limited by Brownian motion. Hum Gene Ther. 1996;7:1527–1534. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
