

Triple-helix formation induces recombination in mammalian cells via a nucleotide excision repair-dependent pathway
- PMID: 10629056
- PMCID: PMC85216
- DOI: 10.1128/MCB.20.3.990-1000.2000
Triple-helix formation induces recombination in mammalian cells via a nucleotide excision repair-dependent pathway
Abstract
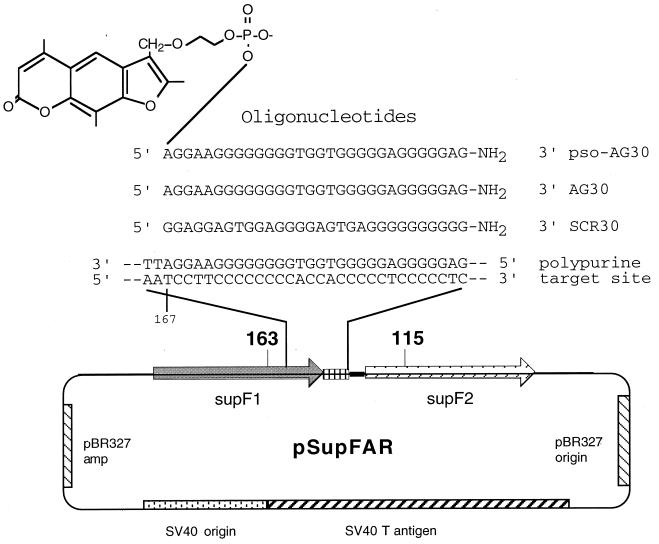
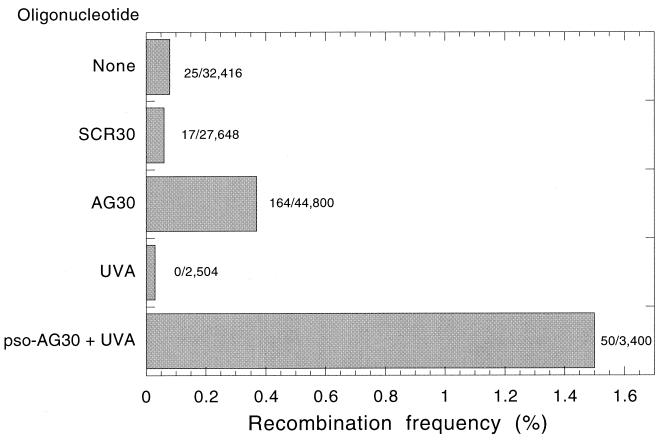
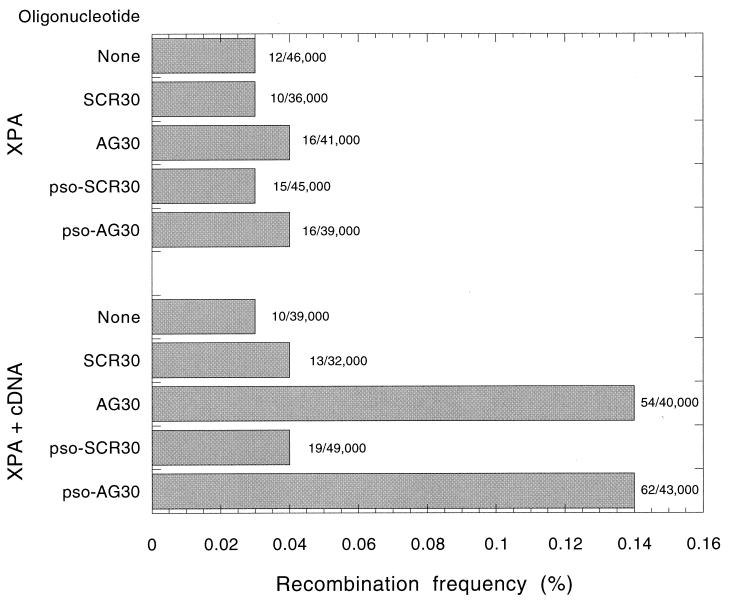
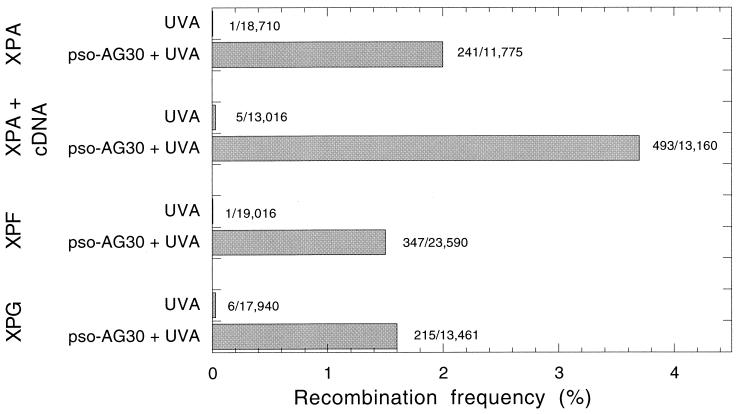
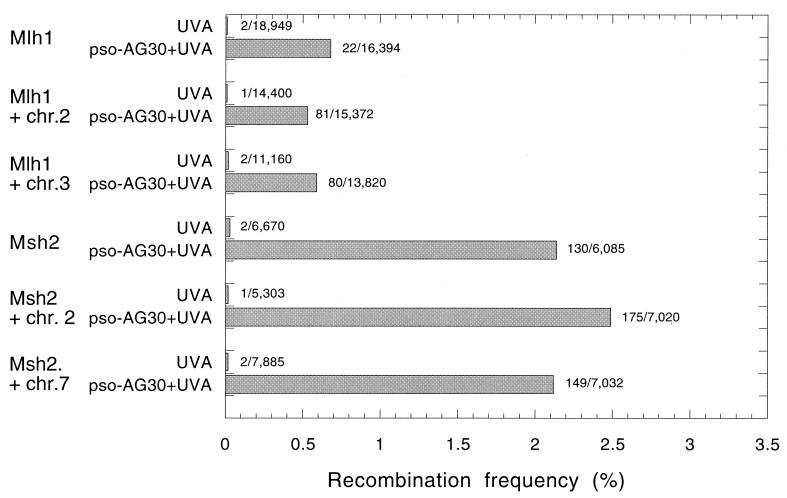
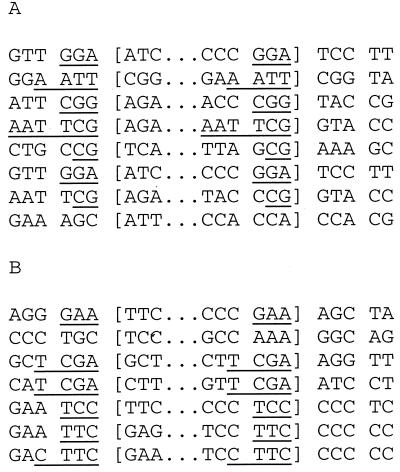
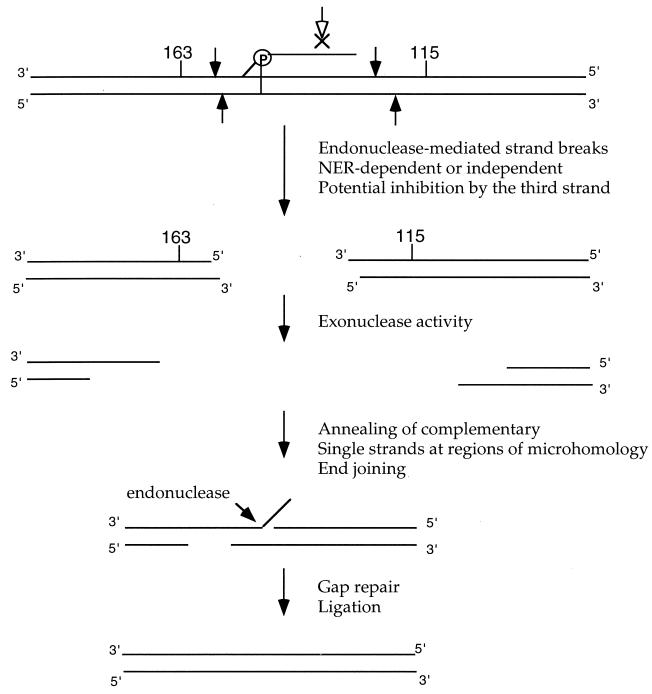
The ability to stimulate recombination in a site-specific manner in mammalian cells may provide a useful tool for gene knockout and a valuable strategy for gene therapy. We previously demonstrated that psoralen adducts targeted by triple-helix-forming oligonucleotides (TFOs) could induce recombination between tandem repeats of a supF reporter gene in a simian virus 40 vector in monkey COS cells. Based on work showing that triple helices, even in the absence of associated psoralen adducts, are able to provoke DNA repair and cause mutations, we asked whether intermolecular triplexes could stimulate recombination. Here, we report that triple-helix formation itself is capable of promoting recombination and that this effect is dependent on a functional nucleotide excision repair (NER) pathway. Transfection of COS cells carrying the dual supF vector with a purine-rich TFO, AG30, designed to bind as a third strand to a region between the two mutant supF genes yielded recombinants at a frequency of 0.37%, fivefold above background, whereas a scrambled sequence control oligomer was ineffective. In human cells deficient in the NER factor XPA, the ability of AG30 to induce recombination was eliminated, but it was restored in a corrected subline expressing the XPA cDNA. In comparison, the ability of triplex-directed psoralen cross-links to induce recombination was only partially reduced in XPA-deficient cells, suggesting that NER is not the only pathway that can metabolize targeted psoralen photoadducts into recombinagenic intermediates. Interestingly, the triplex-induced recombination was unaffected in cells deficient in DNA mismatch repair, challenging our previous model of a heteroduplex intermediate and supporting a model based on end joining. This work demonstrates that oligonucleotide-mediated triplex formation can be recombinagenic, providing the basis for a potential strategy to direct genome modification by using high-affinity DNA binding ligands.
Figures
Similar articles
-
Altered repair of targeted psoralen photoadducts in the context of an oligonucleotide-mediated triple helix.J Biol Chem. 1995 Sep 22;270(38):22595-601. doi: 10.1074/jbc.270.38.22595. J Biol Chem. 1995. PMID: 7673252
-
Targeted correction of an episomal gene in mammalian cells by a short DNA fragment tethered to a triplex-forming oligonucleotide.J Biol Chem. 1999 Apr 23;274(17):11541-8. doi: 10.1074/jbc.274.17.11541. J Biol Chem. 1999. PMID: 10206960
-
Mutagenesis by third-strand-directed psoralen adducts in repair-deficient human cells: high frequency and altered spectrum in a xeroderma pigmentosum variant.Proc Natl Acad Sci U S A. 1996 Apr 2;93(7):2941-6. doi: 10.1073/pnas.93.7.2941. Proc Natl Acad Sci U S A. 1996. PMID: 8610147 Free PMC article.
-
Targeted gene modification using triplex-forming oligonucleotides.Methods Mol Biol. 2004;262:173-94. doi: 10.1385/1-59259-761-0:173. Methods Mol Biol. 2004. PMID: 14769962 Review.
-
Targeted genome modification via triple helix formation.Ann N Y Acad Sci. 2005 Nov;1058:151-61. doi: 10.1196/annals.1359.023. Ann N Y Acad Sci. 2005. PMID: 16394134 Review.
Cited by
-
Replication fork stalling and checkpoint activation by a PKD1 locus mirror repeat polypurine-polypyrimidine (Pu-Py) tract.J Biol Chem. 2012 Sep 28;287(40):33412-23. doi: 10.1074/jbc.M112.402503. Epub 2012 Aug 6. J Biol Chem. 2012. PMID: 22872635 Free PMC article.
-
Human XPA and RPA DNA repair proteins participate in specific recognition of triplex-induced helical distortions.Proc Natl Acad Sci U S A. 2002 Apr 30;99(9):5848-53. doi: 10.1073/pnas.082193799. Epub 2002 Apr 23. Proc Natl Acad Sci U S A. 2002. PMID: 11972036 Free PMC article.
-
Transcription-induced CAG repeat contraction in human cells is mediated in part by transcription-coupled nucleotide excision repair.Mol Cell Biol. 2007 Sep;27(17):6209-17. doi: 10.1128/MCB.00739-07. Epub 2007 Jun 25. Mol Cell Biol. 2007. PMID: 17591697 Free PMC article.
-
Advances in mechanisms of genetic instability related to hereditary neurological diseases.Nucleic Acids Res. 2005 Jul 8;33(12):3785-98. doi: 10.1093/nar/gki697. Print 2005. Nucleic Acids Res. 2005. PMID: 16006624 Free PMC article. Review.
-
Site-directed recombination via bifunctional PNA-DNA conjugates.Proc Natl Acad Sci U S A. 2002 Dec 24;99(26):16695-700. doi: 10.1073/pnas.262556899. Epub 2002 Dec 2. Proc Natl Acad Sci U S A. 2002. PMID: 12461167 Free PMC article.
References
-
- Beal P A, Dervan P B. Second structural motif for recognition of DNA by oligonucleotide-directed triple-helix formation. Science. 1991;251:1360–1363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials