

A physical map, including a BAC/PAC clone contig, of the Williams-Beuren syndrome--deletion region at 7q11.23
- PMID: 10631136
- PMCID: PMC1288354
- DOI: 10.1086/302722
A physical map, including a BAC/PAC clone contig, of the Williams-Beuren syndrome--deletion region at 7q11.23
Abstract
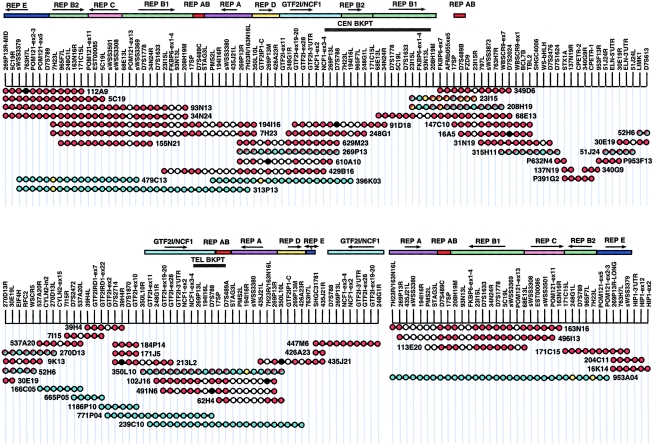
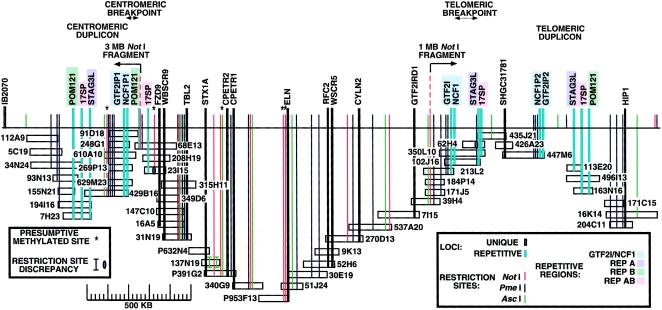
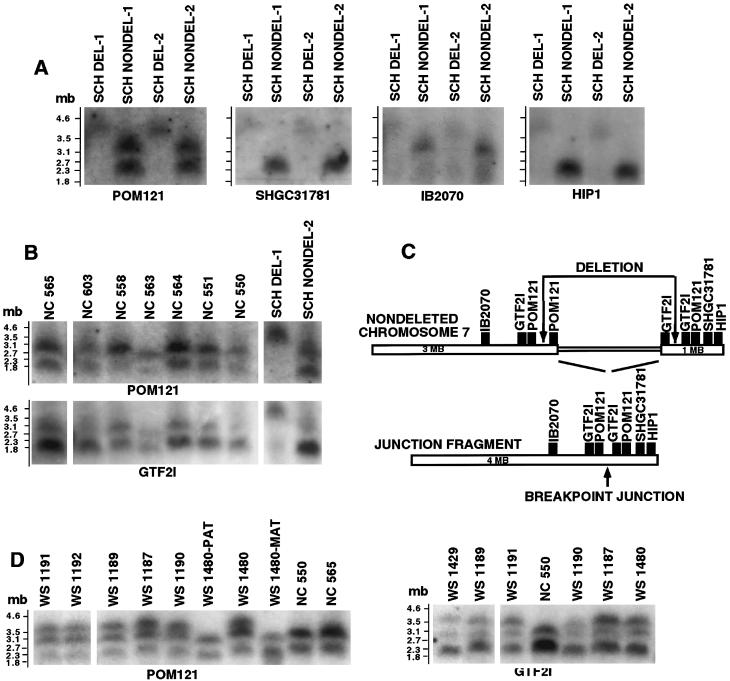
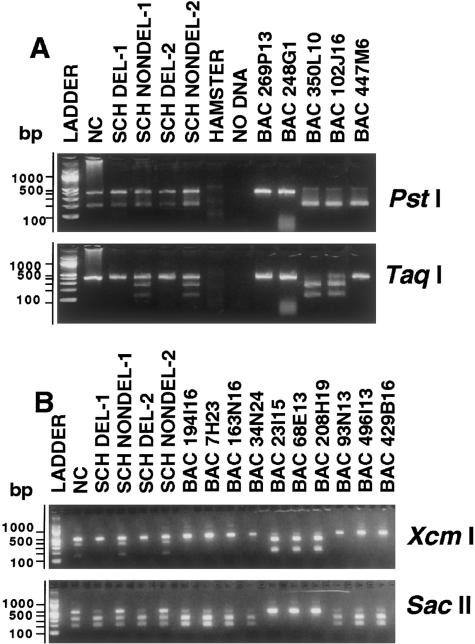
Williams-Beuren syndrome (WBS) is a developmental disorder caused by haploinsufficiency for genes in a 2-cM region of chromosome band 7q11.23. With the exception of vascular stenoses due to deletion of the elastin gene, the various features of WBS have not yet been attributed to specific genes. Although >/=16 genes have been identified within the WBS deletion, completion of a physical map of the region has been difficult because of the large duplicated regions flanking the deletion. We present a physical map of the WBS deletion and flanking regions, based on assembly of a bacterial artificial chromosome/P1-derived artificial chromosome contig, analysis of high-throughput genome-sequence data, and long-range restriction mapping of genomic and cloned DNA by pulsed-field gel electrophoresis. Our map encompasses 3 Mb, including 1.6 Mb within the deletion. Two large duplicons, flanking the deletion, of >/=320 kb contain unique sequence elements from the internal border regions of the deletion, such as sequences from GTF2I (telomeric) and FKBP6 (centromeric). A third copy of this duplicon exists in inverted orientation distal to the telomeric flanking one. These duplicons show stronger sequence conservation with regard to each other than to the presumptive ancestral loci within the common deletion region. Sequence elements originating from beyond 7q11.23 are also present in these duplicons. Although the duplicons are not present in mice, the order of the single-copy genes in the conserved syntenic region of mouse chromosome 5 is inverted relative to the human map. A model is presented for a mechanism of WBS-deletion formation, based on the orientation of duplicons' components relative to each other and to the ancestral elements within the deletion region.
Figures
References
Electronic-Database Information
-
- Genome Sequencing Center, Washington University, St. Louis, http://genome.wustl.edu/gsc/
-
- Human Chromosome 7 Mapping and Sequencing, http://genome.nhgri.nih.gov/chr7/
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for WBS [MIM 194050], CGD [MIM 233700], VCFS [MIM 192430], SMS [MIM 182290], PWS [MIM 176270], AS [MIM 105830], HNPP [MIM 162500], SMAI, II, and III [MIM 253300, MIM 253550, and MIM 253400])
-
- Stanford Human Genome Center, http://www-shgc.stanford.edu/Mapping/index.html
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410 - PubMed
-
- Baumer A, Dutly F, Balmer D, Riegel M, Tukel T, Krajewska-Walasek M, Schinzel AA (1998) High level of unequal meiotic crossovers at the origin of the 22q11.2 and 7q11.23 deletions. Hum Mol Genet 7:887–894 - PubMed
-
- Bouffard GG, Idol JR, Braden VV, Iyer LM, Cunningham AF, Weintraub LA, Touchman JW, et al (1997) A physical map of human chromosome 7: an integrated YAC contig map with average STS spacing of 79 kb. Genome Res 7:673–692 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
