

Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii
- PMID: 10631256
- PMCID: PMC58851
- DOI: 10.1104/pp.122.1.127
Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii
Abstract
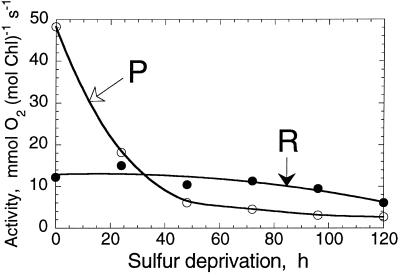
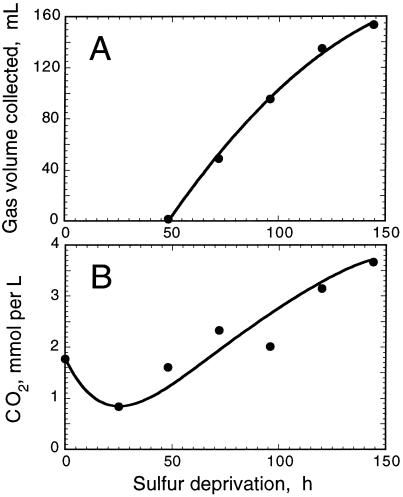
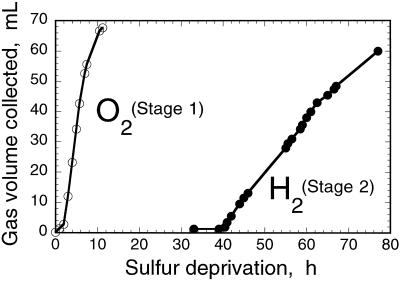
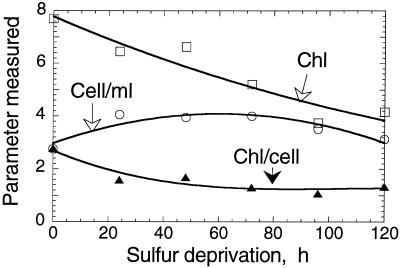
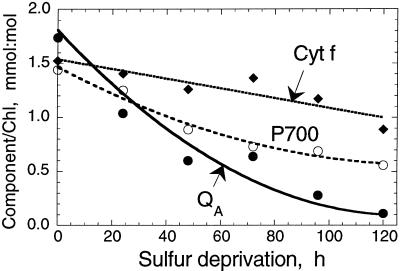
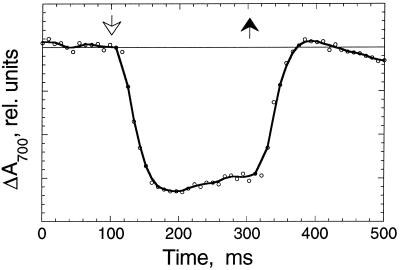
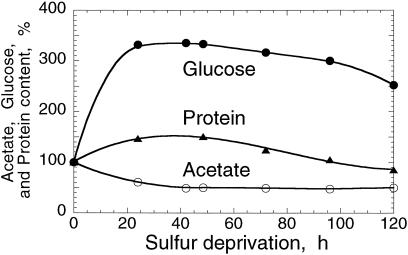
The work describes a novel approach for sustained photobiological production of H(2) gas via the reversible hydrogenase pathway in the green alga Chlamydomonas reinhardtii. This single-organism, two-stage H(2) production method circumvents the severe O(2) sensitivity of the reversible hydrogenase by temporally separating photosynthetic O(2) evolution and carbon accumulation (stage 1) from the consumption of cellular metabolites and concomitant H(2) production (stage 2). A transition from stage 1 to stage 2 was effected upon S deprivation of the culture, which reversibly inactivated photosystem II (PSII) and O(2) evolution. Under these conditions, oxidative respiration by the cells in the light depleted O(2) and caused anaerobiosis in the culture, which was necessary and sufficient for the induction of the reversible hydrogenase. Subsequently, sustained cellular H(2) gas production was observed in the light but not in the dark. The mechanism of H(2) production entailed protein consumption and electron transport from endogenous substrate to the cytochrome b(6)-f and PSI complexes in the chloroplast thylakoids. Light absorption by PSI was required for H(2) evolution, suggesting that photoreduction of ferredoxin is followed by electron donation to the reversible hydrogenase. The latter catalyzes the reduction of protons to molecular H(2) in the chloroplast stroma.
Figures
References
-
- Adams MWW. The structure and mechanism of iron-hydrogenases. Biochim Biophys Acta. 1990;1020:115–145. - PubMed
-
- Albracht SPJ. Nickel hydrogenases: in search of the active site. Biochim Biophys Acta. 1994;1188:167–204. - PubMed
-
- Aoyama K, Uemura I, Miyake J, Asada Y. Fermentative metabolism to produce hydrogen gas and organic compounds in a cyanobacterium, Spirulina platensis. J Ferment Bioenerg. 1997;83:17–20.
-
- Appel J, Schulz R. Hydrogen metabolism in organisms with oxygenic photosynthesis: hydrogenases as important regulatory devices for a proper redox poising? J Photochem Photobiol. 1998;47:1–11.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
