

A highly conserved enhancer in the Dlx5/Dlx6 intergenic region is the site of cross-regulatory interactions between Dlx genes in the embryonic forebrain
- PMID: 10632600
- PMCID: PMC6772408
- DOI: 10.1523/JNEUROSCI.20-02-00709.2000
A highly conserved enhancer in the Dlx5/Dlx6 intergenic region is the site of cross-regulatory interactions between Dlx genes in the embryonic forebrain
Abstract
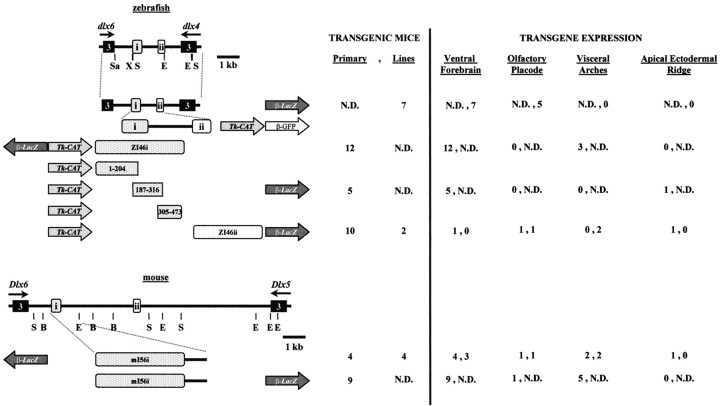
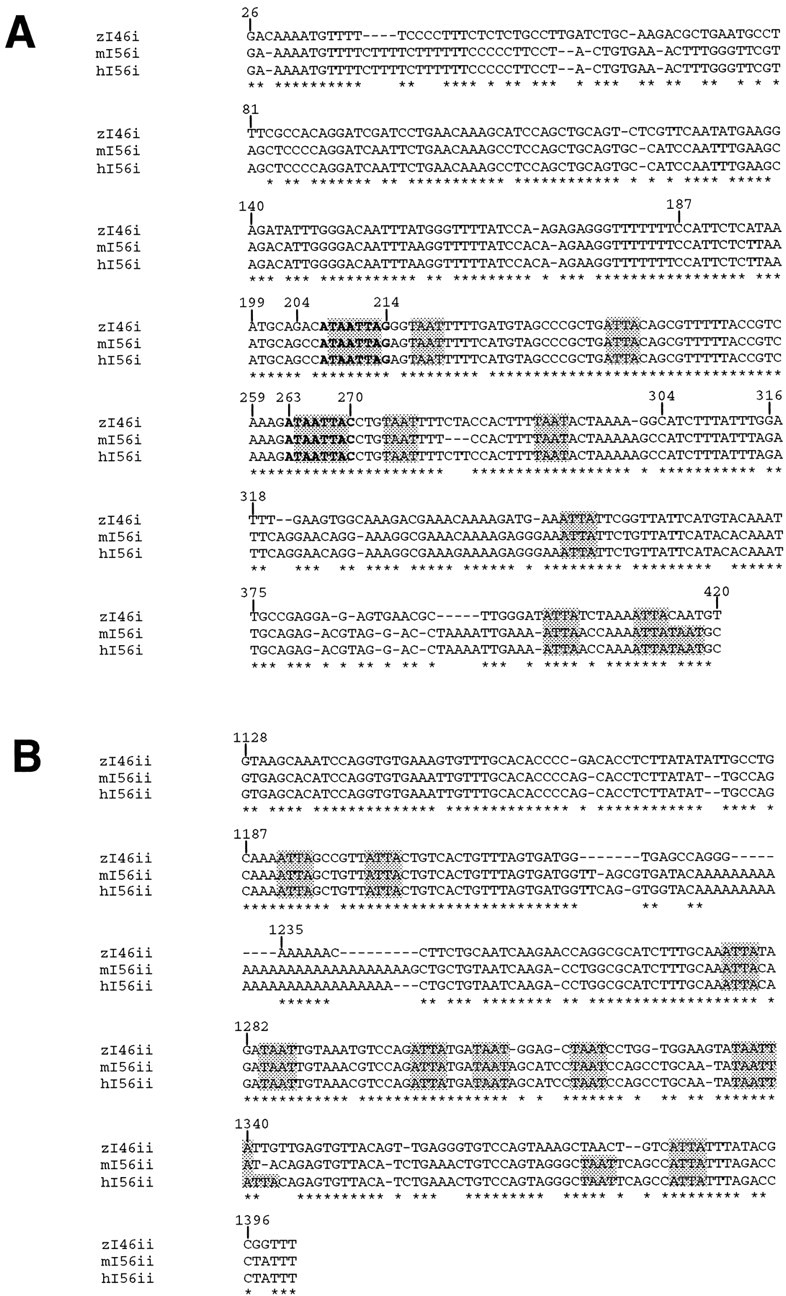
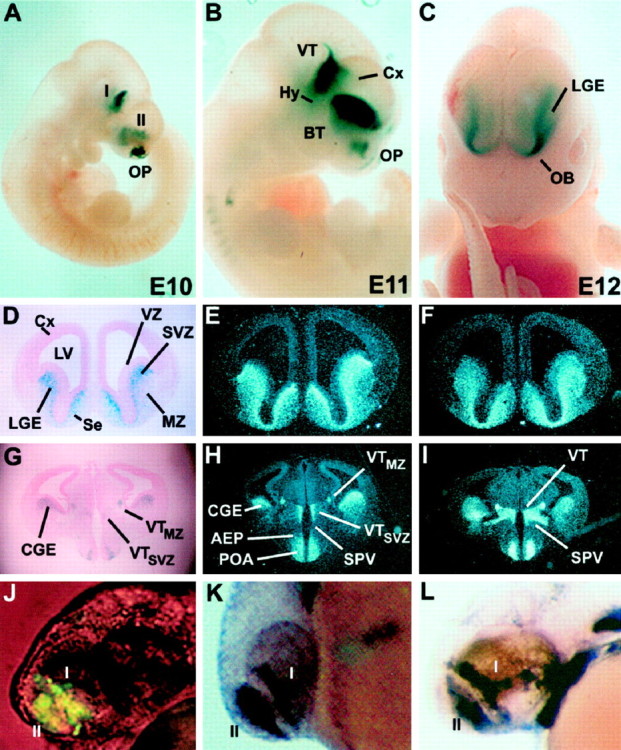
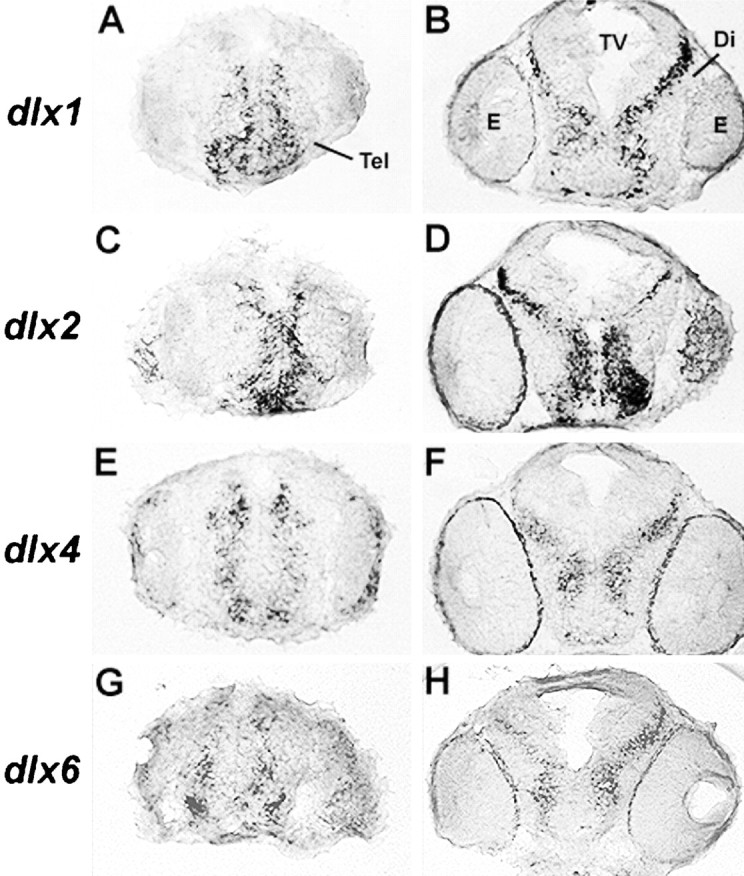
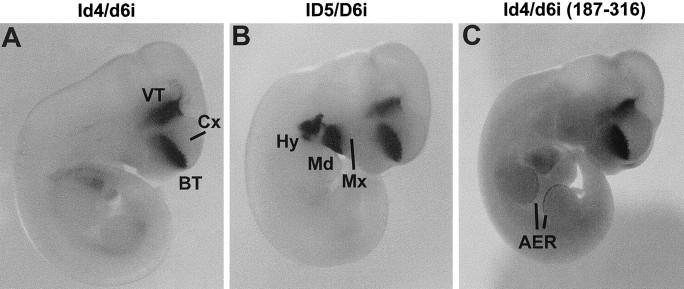
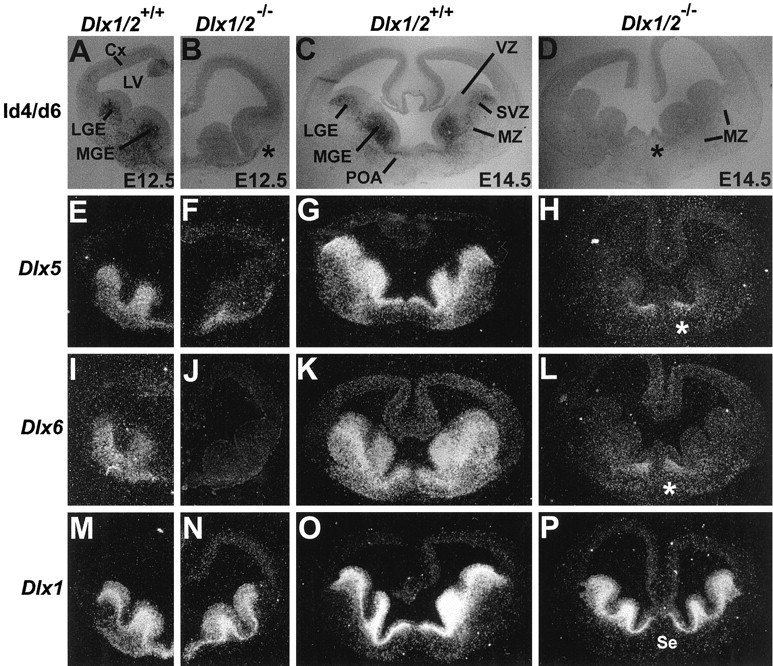
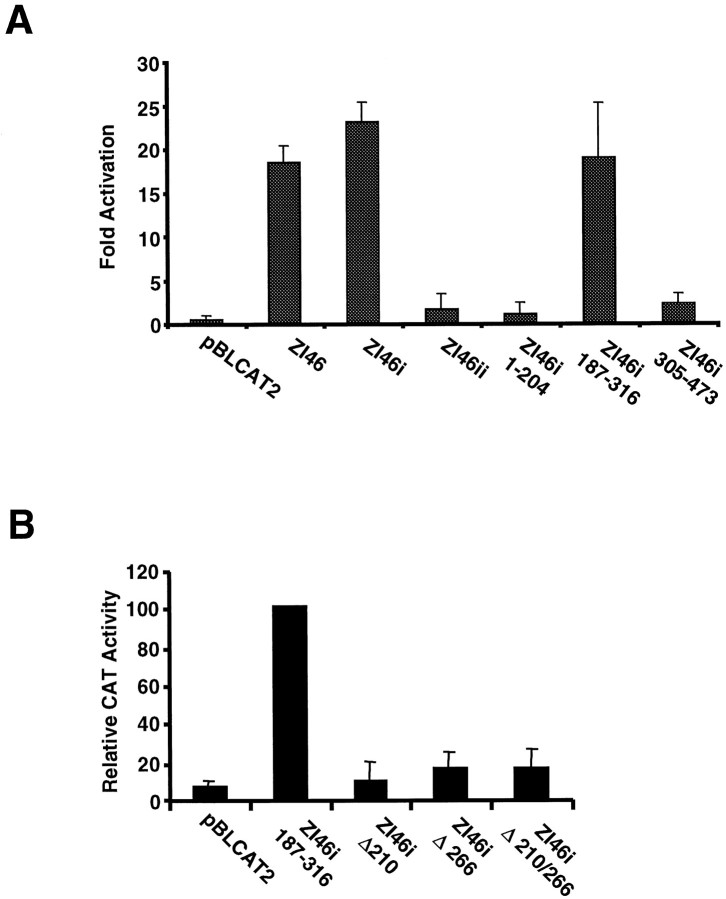
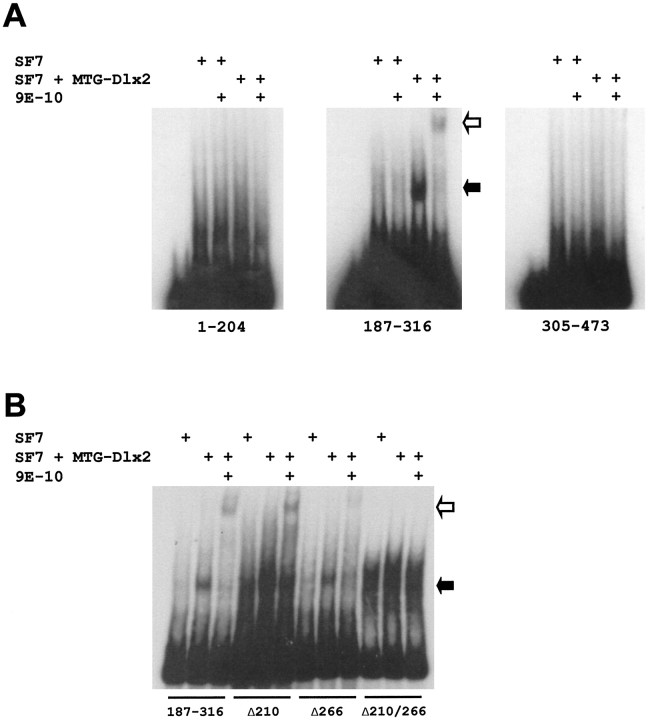
Four Dlx homeobox genes, Dlx1, Dlx2, Dlx5, and Dlx6 are expressed in the same primordia of the mouse forebrain with temporally overlapping patterns. The four genes are organized as two tail-to-tail pairs, Dlx1/Dlx2 and Dlx5/Dlx6, a genomic arrangement conserved in distantly related vertebrates like zebrafish. The Dlx5/Dlx6 intergenic region contains two sequences of a few hundred base pairs, remarkably well conserved between mouse and zebrafish. Reporter transgenes containing these two sequences are expressed in the forebrain of transgenic mice and zebrafish with patterns highly similar to endogenous Dlx5 and Dlx6 expression. The activity of the transgene is drastically reduced in mouse mutants lacking both Dlx1 and Dlx2, consistent with the decrease in endogenous Dlx5 and Dlx6 expression. These results suggest that cross-regulation by Dlx proteins, mediated by the intergenic sequences, is essential for Dlx5 and Dlx6 expression in the forebrain. This hypothesis is supported by cotransfection and DNA-protein binding experiments. We propose that the Dlx genes are part of a highly conserved developmental pathway that regulates forebrain development.
Figures
References
-
- Acampora D, Merlo GR, Paleari L, Zeraga B, Postiglione MP, Mantero S, Bober E, Barbieri O, Simeone A, Levi G. Craniofacial, vestibular and bone defects in mice lacking the Distal-less-related gene Dlx5. Development. 1999;126:3795–3809. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JLR. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997a;278:474–476. - PubMed
-
- Anderson SA, Qiu M, Bulfone A, Eisenstat DD, Meneses J, Pedersen R, Rubenstein JLR. Mutations of the homeobox genes Dlx-1 and Dlx-2 disrupt the striatal subventricular zone and differentiation of late-born striatal neurons. Neuron. 1997b;19:27–37. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials