

Osmotic stress induces rapid activation of a salicylic acid-induced protein kinase and a homolog of protein kinase ASK1 in tobacco cells
- PMID: 10634915
- PMCID: PMC149182
Osmotic stress induces rapid activation of a salicylic acid-induced protein kinase and a homolog of protein kinase ASK1 in tobacco cells
Erratum in
- Plant Cell 2000 Apr;12(4):611
Abstract
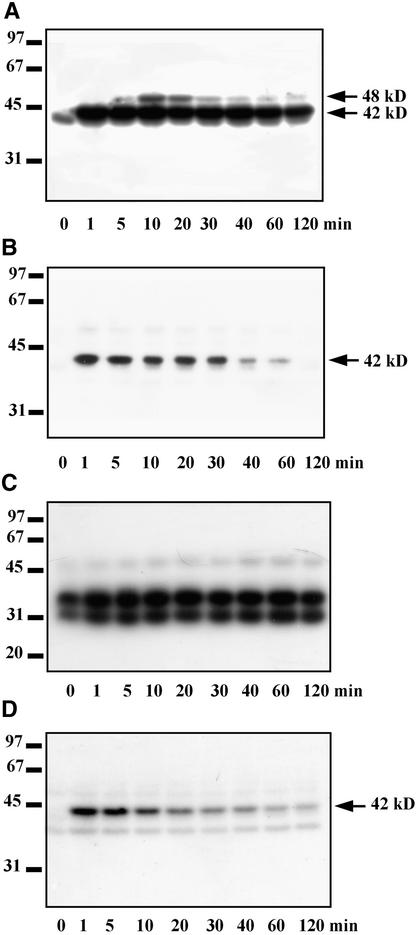
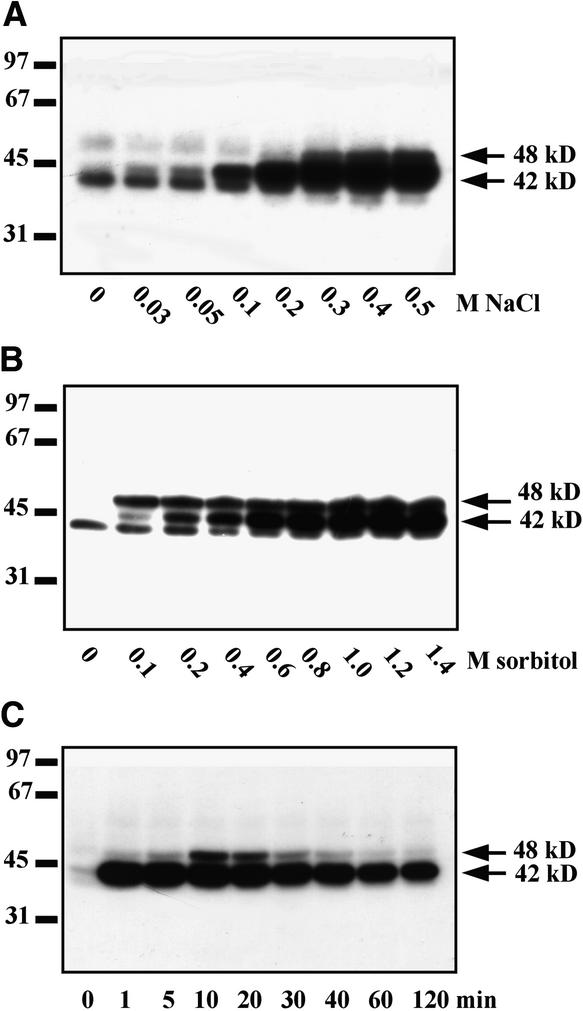
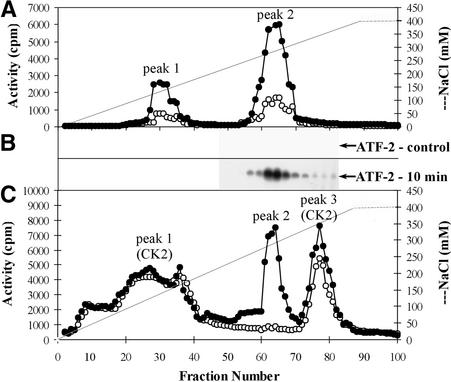
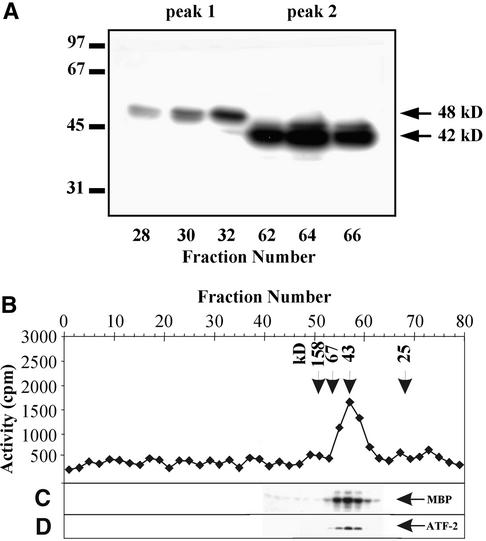
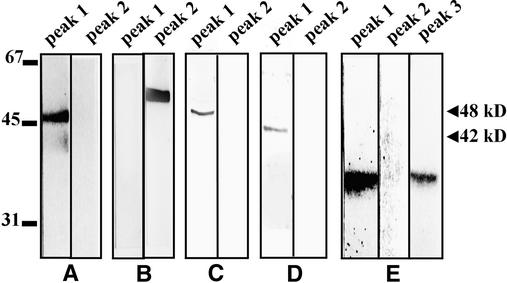
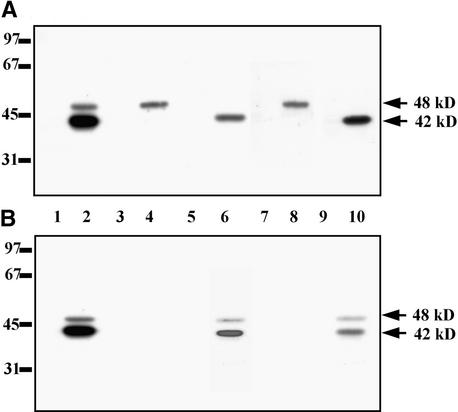
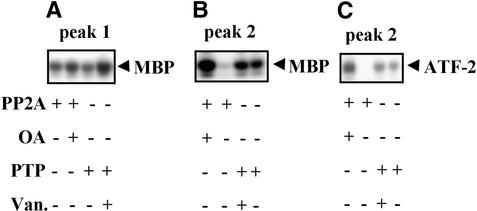
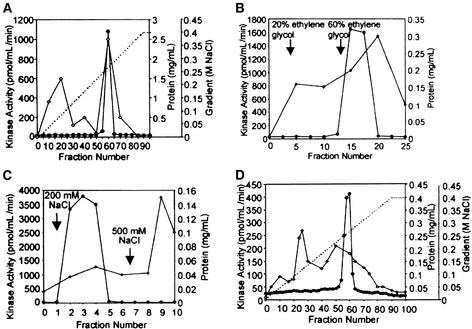
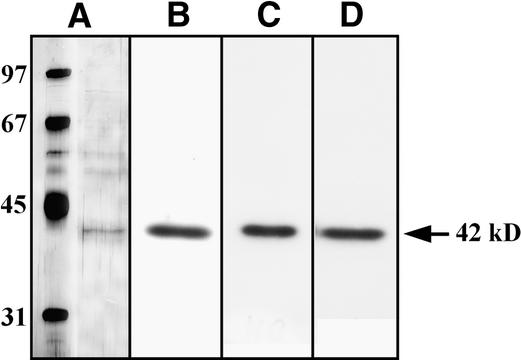
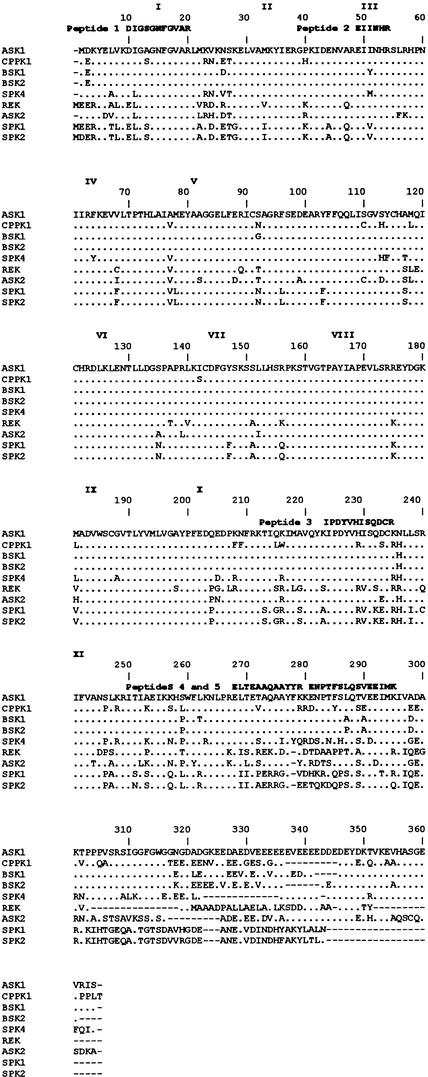
In tobacco cells, osmotic stress induced the rapid activation of two protein kinases that phosphorylate myelin basic protein. Immunological studies demonstrated that the 48-kD kinase is the salicylic acid-induced protein kinase (SIPK), a member of the mitogen-activated protein kinase family. SIPK was activated 5 to 10 min after the cells were exposed to osmotic stresses, and its activity persisted for approximately 30 min. In contrast, the 42-kD kinase was activated within 1 min after osmotic stress, and its activity was maintained for approximately 2 hr. Moreover, in addition to myelin basic protein, the 42-kD kinase phosphorylated casein and two transcription factors, c-Jun and ATF-2. This latter enzyme was inactivated by a serine/threonine-specific phosphatase but, unlike SIPK, was not affected by a tyrosine-specific phosphatase. After the 42-kD kinase was purified to apparent homogeneity, tryptic peptide analysis indicated that it is a homolog of Arabidopsis serine/threonine kinase1 (ASK1).
Figures
References
-
- Bögre, L., Ligterink, W., Heberle-Bors, E., and Hirt, H. (1996). Mechanosensors in plants. Nature 383 489–490. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous