

doi: 10.1093/emboj/19.2.273.
A nuclear tyrosine phosphorylation circuit: c-Jun as an activator and substrate of c-Abl and JNK
Affiliations
- PMID: 10637231
- PMCID: PMC305561
- DOI: 10.1093/emboj/19.2.273
Item in Clipboard
A nuclear tyrosine phosphorylation circuit: c-Jun as an activator and substrate of c-Abl and JNK
EMBO J.
.
Abstract
The nuclear function of the c-Abl tyrosine kinase is not well understood. In order to identify nuclear substrates of Abl, we constructed a constitutively active and nuclear form of the protein. We found that active nuclear Abl efficiently phosphorylate c-Jun, a transcription factor not previously known to be tyrosine phosphorylated. After phosphorylation of c-Jun by Abl on Tyr170, both proteins interacted via the SH2 domain of Abl. Surprisingly, elevated levels of c-Jun activated nuclear Abl, resulting in activation of the JNK serine/threonine kinase. This phosphorylation circuit generates nuclear tyrosine phosphorylation and represents a reversal of previously known signalling models.
Figures
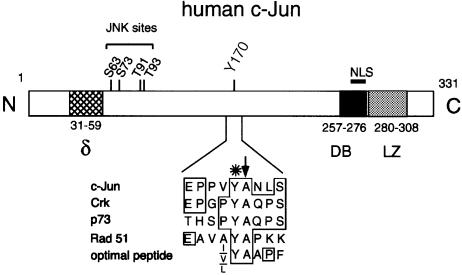
Fig. 1. Cartoon of the human c–Jun primary structure and domain arrangement. The sequence surrounding Tyr170 (residues 166–174) is shown in an enlargement. An asterisk marks the tyrosine that is a site of putative phosphorylation. A vertical arrow indicates the alanine residue following the tyrosine that is typical of good Abl substrates. These are human c-Crk, Tyr221 (Feller et al., 1994); human p73, Tyr99 (Yuan et al., 1999); human Rad51, Tyr54 (Yuan et al., 1998); and optimal peptide (Songyang et al., 1995). The following regions are highlighted: the delta domain (δ), the DNA-binding domain (DB) and the leucine zipper dimerization domain (LZ). The location of the NLS and of the N–terminal phosphorylation sites (JNK sites) is shown. N, N–terminus; C, C–terminus.
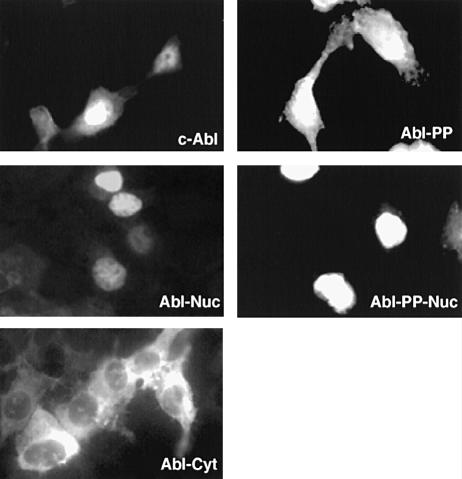
Fig. 2. Subcellular localization of various Abl forms by immuno- fluorescence. HEK-293 cells were plated onto glass coverslips 16 h before transfection with several forms of Abl (5 μg) as shown in each panel. Cells were fixed 48 h after transfection. The Abl proteins were stained with mouse monoclonal anti–Abl antibody.
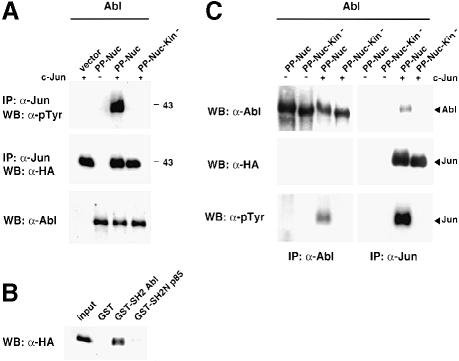
Fig. 3. Abl phosphorylates and interacts with Jun. (A) Tyrosine phosphorylation of Jun. HEK-293 cells were transiently transfected with CMV-HA-c–Jun (5 μg) and CMV–Abl-PP-Nuc or CMV–Abl-PP-Nuc-Kin– (2 μg) as a control. A 500 μg aliquot of total protein extract was immunoprecipitated with anti–Jun antibodies. Immunoblots were probed with anti-phosphotyrosine antibodies (upper panel) or antibodies against HA (middle panel). Abl protein levels are shown in the lower panel. (B) GST binding. HEK-293 cells were transiently transfected with CMV–Abl-PP-Nuc (2 μg) and CMV-HA-c–Jun (5 μg). A 500 μg aliquot of total protein extract was incubated in the presence of the various GST fusions pre-bound to glutathione beads. Pull-downs were analysed by immunoblotting. c–Jun was revealed using antibodies against the HA tag. (C) Co-immunoprecipitation of Abl and Jun from transiently transfected HEK-293 cells. Cells were transfected with CMV-HA-c–Jun (5 μg) and CMV–Abl-PP-Nuc (2 μg), or its catalytically inactive version, CMV–Abl-PP-Nuc-Kin–, as a control. A 1 mg aliquot of total protein extract was immunoprecipitated with antibodies against Abl (left panels) or with the anti–Jun antibody (right panels). The membranes were probed with anti–Abl antibodies (upper panels), anti-HA antibodies (middle panels) and anti-phosphotyrosine antibodies (lower panels). Migration of the Abl and Jun proteins is indicated by the arrows.
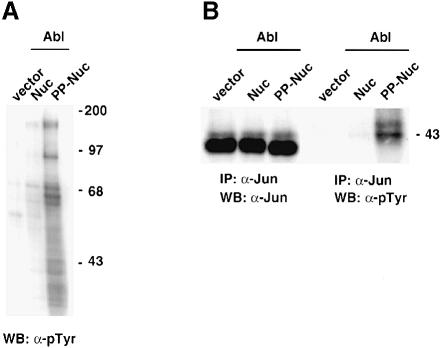
Fig. 4. Phosphorylation of endogenous Jun. (A) Abl can induce phosphorylation of several proteins in the nucleus. NIH-3T3 cells were transfected with CMV–Abl-Nuc or CMV–Abl-PP-Nuc (5 μg). A 50 μg aliquot of nuclear protein extracts was separated, blotted and probed with anti-phosphotyrosine antibodies. (B) Abl can induce phosphorylation of endogenous c–Jun. NIH-3T3 cells were transfected with CMV–Abl-Nuc or CMV–Abl-PP-Nuc (5 μg). A 500 μg aliquot of nuclear protein extract was immunoprecipitated with anti-c–Jun antibodies. Blots of immunoprecipitates were probed with antibodies against Jun (left panel) or anti-phosphotyrosine (right panel).
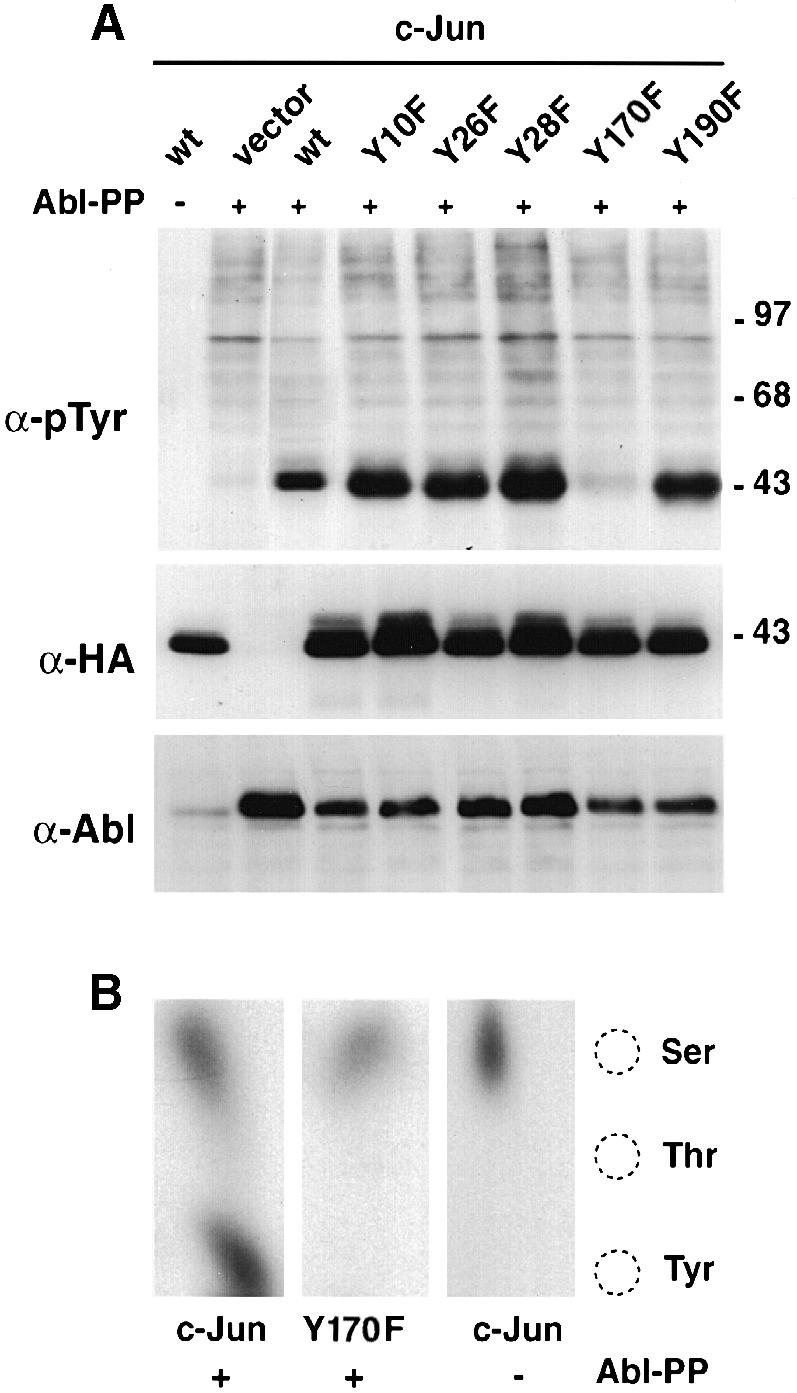
Fig. 5. Mapping of the tyrosine phosphorylation site in c–Jun. (A) HEK-293 cells were transfected with c–Jun tyrosine mutants (CMV-HA constructs, 5 μg) and pSGT–Abl-PP (5 μg). Total protein extracts were separated, blotted and probed with anti-phosphotyrosine (top panel), anti-HA (middle panel) and anti–Abl antibodies (lower panel). (B) In vivo labelling. CMV-HA-c–Jun or CMV-HA–Jun-Y170F (5 μg) were transfected in HEK-293 cells along with pSGT–Abl-PP (5 μg) in the presence of [32P]orthophosphate. Total protein extracts were immunoprecipitated with anti–Jun antibodies, separated and blotted. The band corresponding to Jun was revealed by auto- radiography, excised and subjected to phosphoamino acid analysis using thin-layer chromatography. Migration of the phosphoaminoacid markers is shown on the right.
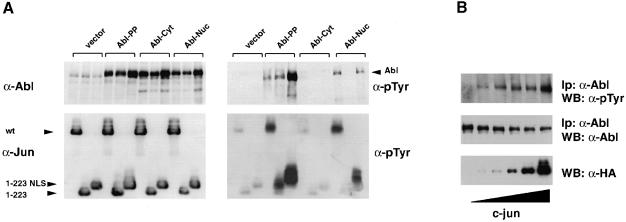
Fig. 6. Jun is a substrate and an activator of Abl in the nucleus. (A) HEK-293 cells were transiently co-transfected with various Abl and Jun forms (5 μg each). Abl forms were: Abl-PP, bearing the activating double proline mutation; Abl-Cyt, mutated in the NLS; and Abl-Nuc, mutated in the NES. Jun forms were: wild-type c–Jun; Jun1–223, a C–terminal truncation lacking the DNA-binding, NLS and leucine zipper domains; and Jun1–223-NLS, identical to the previous form but bearing an added NLS from the SV40 large T antigen. A 50 μg aliquot of total protein extract was processed for immunoblotting. Samples were separated by 7.5% (Abl) and 10% (Jun) SDS–PAGE. Left panels: protein levels as indicated. Right panels: phosphotyrosine levels. Migration of the Abl and Jun proteins is indicated by the arrows. (B) HEK-293 cells were transiently transfected with Abl-Nuc (5 μg) and increasing amounts of c–Jun (0.25, 0.5, 1, 2 and 4 μg). A 50 μg aliquot of total protein extract was processed for immunoblotting with anti-HA antibodies (bottom panel). Abl activation was measured by anti-phosphotyrosine immunoblotting after immunoprecipitation (top panel). Levels of immunoprecipitated Abl are shown below (middle panel).
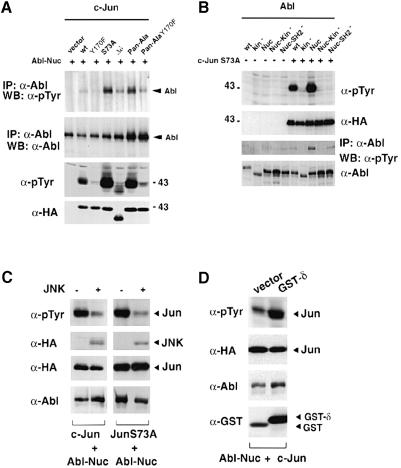
Fig. 7. Structural requirements for Abl activation. (A) HEK-293 cells were transiently transfected with CMV–Abl-Nuc (5 μg) and different Jun mutants (5 μg each). Abl activation was measured by anti-phosphotyrosine immunoblotting after immunoprecipitation (top panel). Levels of immunoprecipitated Abl are shown below. Levels and the tyrosine phosphorylation of the different Jun proteins were detected using anti-HA antibodies or anti-phosphotyrosine antibodies, respectively (lower two panels). (B) Ability of several Abl mutants to phosphorylate Jun. HEK-293 cells were transiently transfected with various Abl forms (5 μg) as indicated, with our without CMV-HA-c–Jun-S73A (5 μg). Lysates were analysed by immunoblotting using specific antibodies as indicated. (C) JNK negatively affects Jun phosphorylation by Abl. HEK-293 cells were transfected with CMV–Abl-Nuc (2 μg) and CMV-HA-c–Jun or CMV-HA-c–JunS73A (2 μg) in the presence or absence of SV40-HA-JNK (6 μg). Total protein was immunoblotted and probed with anti-phosphotyrosine antibodies (top panel), anti-HA antibodies (HA–Jun and HA-JNK, two central panels) or by anti–Abl antibodies (bottom panel). (D) Expression of the Jun δ region enhances the ability of Jun to be phosphorylated by Abl. HEK-293 cells were transfected with CMV–Abl-Nuc (2 μg) and CMV-HA-c–Jun (2 μg) in the presence of pEBG or pEBG-δ–Jun (6 μg). Phosphorylation of c–Jun protein was measured by anti-phosphotyrosine antibodies (top panel) and protein levels were detected by specific antibodies (lower panels).
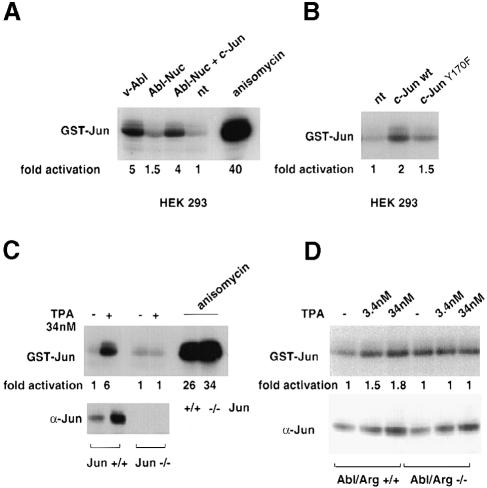
Fig. 8. Activation of JNK by Abl and Jun. (A) JNK kinase assay on GST–Jun pull-downs from HEK-293 cells transfected with nuclear Abl (CMV–Abl-Nuc, 5 μg) in the presence of CMV-HA-c–Jun or control vector (5 μg). Positive controls were extracts from cells transfected with pSGT-v–Abl (5 μg) or cells treated with 0.5 μg/ml anisomycin for 40 min. Total proteins were incubated with ∼10 μg of GST–Jun1–101 pre-bound to glutathione beads. After the kinase reaction, phosphorylation was analysed by SDS–PAGE and revealed by autoradiography. To calculate JNK activation, bands were excised and incorporated radioactivity measured by scintillation counting. The values represent the average from at least two independent experiments. (B) JNK kinase assay on GST–Jun pull-downs from HEK-293 cells transfected with CMV-HA-c–Jun or with CMV-HA-c–JunY170F (10 μg) was performed as described in (A). (C) Jun+/+ and Jun–/– mouse fibroblasts (Schreiber et al., 1999) were kept in normal serum and treated with 34 nM TPA for 14 h or with 0.5 μg/ml anisomycin for 40 min. Jun levels in nuclear extracts were detected by immunoblotting, and the JNK kinase assay was performed as described above. (D) Abl/Arg+/+ and Abl/Arg–/– mouse embryo (Koleske et al., 1998) fibroblasts were kept in normal serum treated with increasing amounts of TPA for 14 h. Detection of Jun levels and JNK kinase assay were performed as described above.
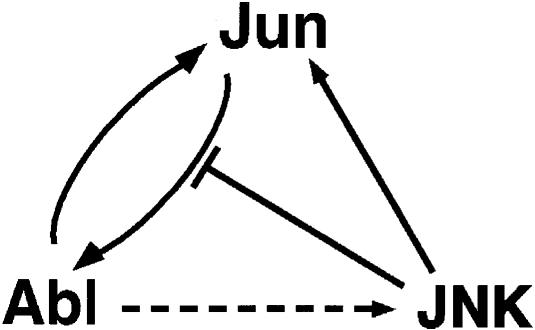
Fig. 9. Diagram illustrating the possible functional relationship between Jun, Abl and JNK. Elevated Jun levels cause an increase in the activity of nuclear Abl, possibly by a direct mechanism (solid forward arrow). Abl phosphorylates Jun on tyrosine (solid backward arrow), increasing the ability of Jun to activate Abl further (positive feedback loop). Abl activity results in an increase in JNK activity, most likely by an indirect mechanism (dashed arrow). Active JNK in turn phosphorylates Jun (solid arrow). Elevated JNK levels negatively affect Jun phosphorylation by Abl (blunt arrow).
References
-
- Agami R., Blandino, G., Oren, M. and Shaul, Y. (1999) Interaction of c–Abl and p73α and their collaboration to induce apoptosis. Nature, 399, 809–813. - PubMed
-
- Angel P., Smeal, T., Meek, J. and Karin, M. (1989) Jun and v-jun contain multiple regions that participate in transcriptional activation in an interdependent manner. New Biol., 1, 35–43. - PubMed
-
- Baichwal V.R. and Tjian, R. (1990) Control of c–Jun activity by interaction of a cell-specific inhibitor with regulatory domain δ: differences between v- and c–Jun. Cell, 63, 815–825. - PubMed
-
- Bannister A.J., Oehler, T., Wilhelm, D., Angel, P. and Kouzarides, T. (1995) Stimulation of c–Jun activity by CBP: c–Jun residues Ser63/73 are required for CBP induced stimulation in vivo and CBP binding in vitro. Oncogene, 11, 2509–2514. - PubMed
-
- Barilá D. and Superti-Furga, G. (1998) An intramolecular SH3-domain interaction regulates c–Abl activity. Nature Genet., 18, 280–282. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
