

Regulation of mitotic inhibitor Mik1 helps to enforce the DNA damage checkpoint
- PMID: 10637286
- PMCID: PMC14752
- DOI: 10.1091/mbc.11.1.1
Regulation of mitotic inhibitor Mik1 helps to enforce the DNA damage checkpoint
Abstract
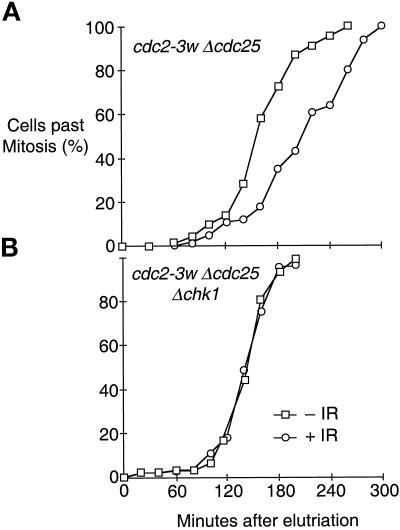
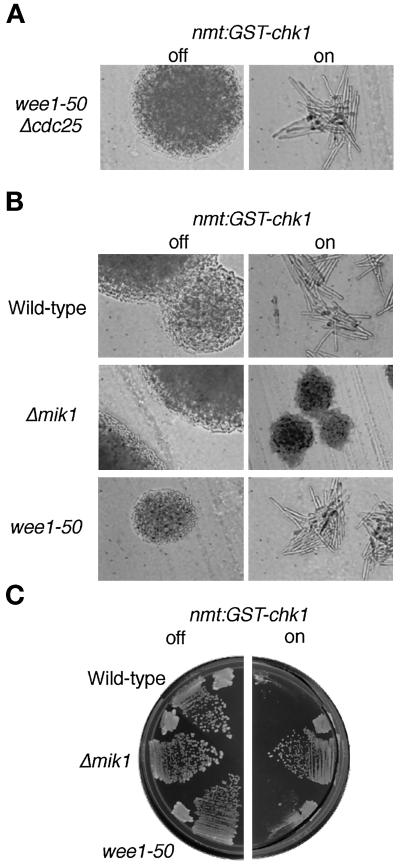
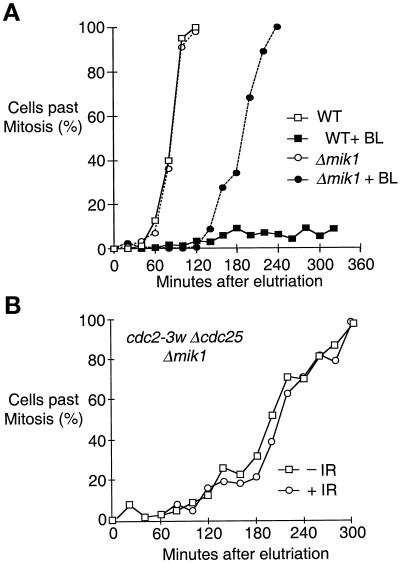
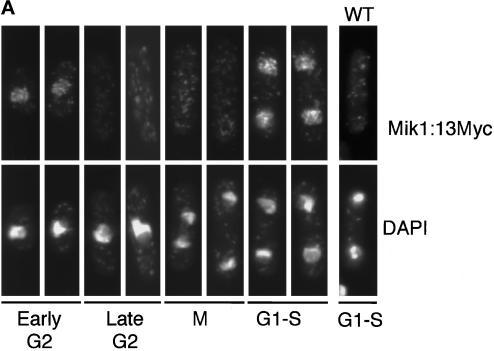
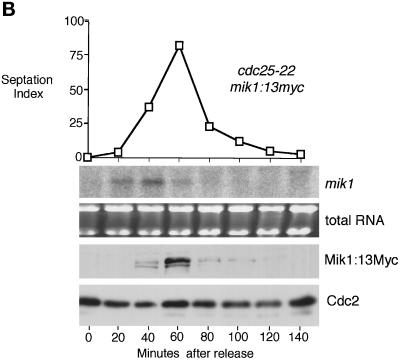
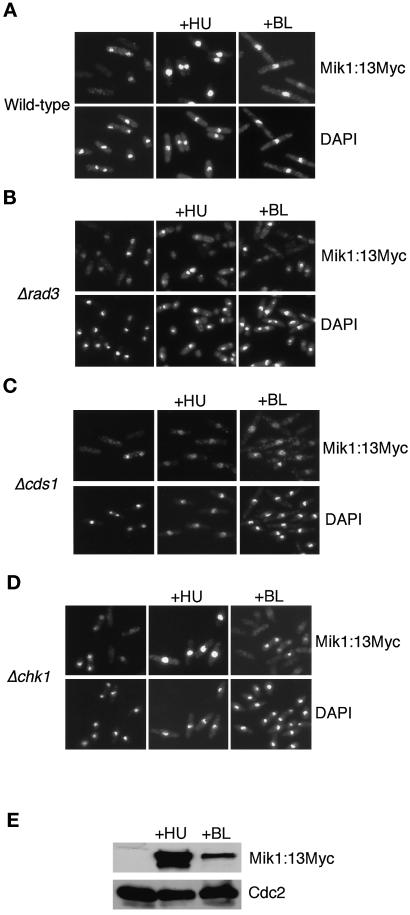
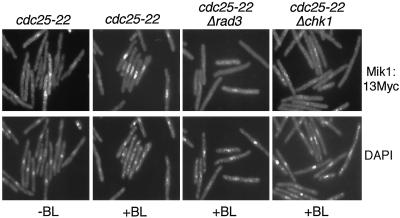
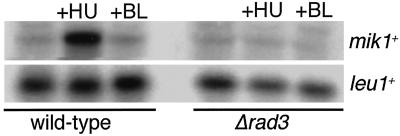
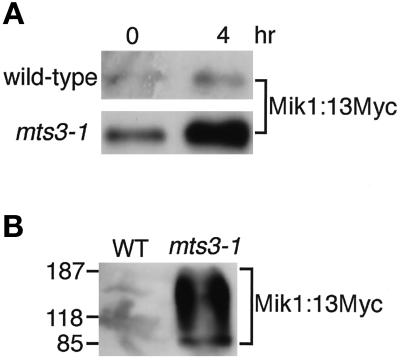
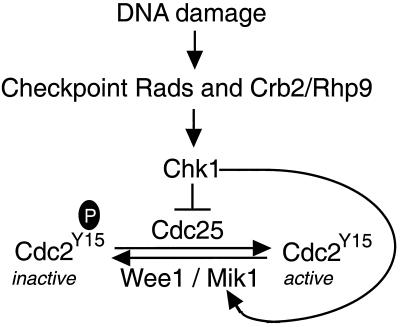
The protein kinase Chk1 enforces the DNA damage checkpoint. This checkpoint delays mitosis until damaged DNA is repaired. Chk1 regulates the activity and localization of Cdc25, the tyrosine phosphatase that activates the cdk Cdc2. Here we report that Mik1, a tyrosine kinase that inhibits Cdc2, is positively regulated by the DNA damage checkpoint. Mik1 is required for checkpoint response in strains that lack Cdc25. Long-term DNA damage checkpoint arrest fails in Deltamik1 cells. DNA damage increases Mik1 abundance in a Chk1-dependent manner. Ubiquitinated Mik1 accumulates in a proteasome mutant, which indicates that Mik1 normally has a short half-life. Thus, the DNA damage checkpoint might regulate Mik1 degradation. Mik1 protein and mRNA oscillate during the unperturbed cell cycle, with peak amounts detected around S phase. These data indicate that regulation of Mik1 abundance helps to couple mitotic onset to the completion of DNA replication and repair. Coordinated negative regulation of Cdc25 and positive regulation of Mik1 ensure the effective operation of the DNA damage checkpoint.
Figures
References
-
- Blasina A, Van de Weyer I, Laus MC, Luyten WHML, Parker AE, McGowan CH. A human homolog of the checkpoint kinase Cds1 directly inhibits Cdc25. Curr Biol. 1999;9:1–10. - PubMed
-
- Boddy MN, Furnari B, Mondesert O, Russell P. Replication checkpoint enforced by kinases Cds1 and Chk1. Science. 1998;280:909–912. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
