

Modulation of CICR has no maintained effect on systolic Ca2+: simultaneous measurements of sarcoplasmic reticulum and sarcolemmal Ca2+ fluxes in rat ventricular myocytes
- PMID: 10639102
- PMCID: PMC2269754
- DOI: 10.1111/j.1469-7793.2000.t01-2-00259.x
Modulation of CICR has no maintained effect on systolic Ca2+: simultaneous measurements of sarcoplasmic reticulum and sarcolemmal Ca2+ fluxes in rat ventricular myocytes
Abstract
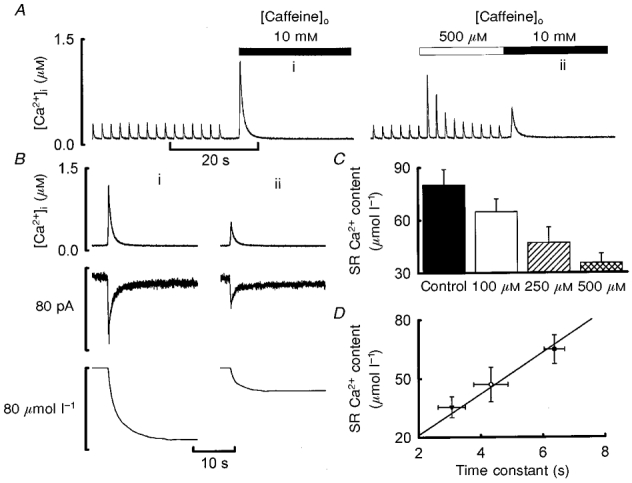
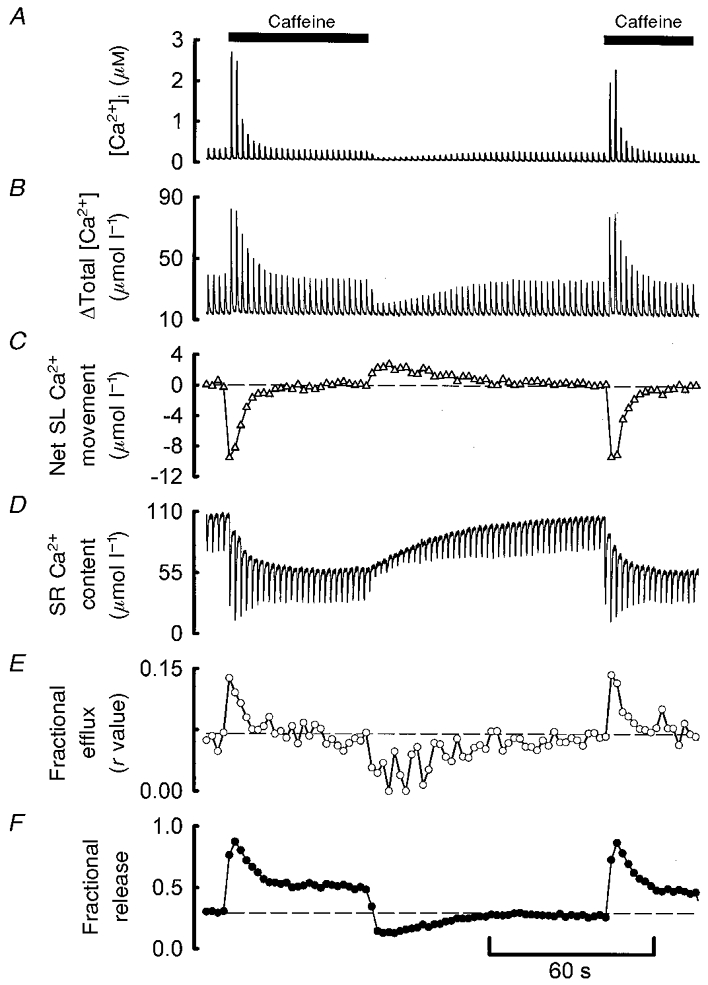
1. The effects of modulating Ca2+-induced Ca2+ release (CICR) in single cardiac myocytes were investigated using low concentrations of caffeine (< 500 microM) in reduced external Ca2+ (0.5 mM). Caffeine produced a transient potentiation of systolic [Ca2+]i (to 800 % of control) which decayed back to control levels. 2. Caffeine decreased the steady-state sarcoplasmic reticulum (SR) Ca2+ content. As the concentration of caffeine was increased, both the potentiation of the systolic Ca2+ transient and the decrease in SR Ca2+ content were increased. At higher concentrations, the potentiating effect decayed more rapidly but the rate of recovery on removal of caffeine was unaffected. 3. A simple model in which caffeine produces a fixed increase in the fraction of SR Ca2+ which is released could account qualitatively but not quantitatively for the above results. 4. The changes in total [Ca2+] during systole were obtained using measurements of the intracellular Ca2+ buffering power. Caffeine initially increased the fractional release of SR Ca2+. This was followed by a decrease to a level greater than that under control conditions. The fraction of systolic Ca2+ which was pumped out of the cell increased abruptly upon caffeine application but then recovered back to control levels. The increase in fractional loss is due to the fact that, as the cytoplasmic buffers become saturated, a given increase in systolic total [Ca2+] produces a larger increase in free [Ca2+] and thence of Ca2+ efflux. 5. These results confirm that modulation of the ryanodine receptor has no maintained effect on systolic Ca2+ and show the interdependence of SR Ca2+ content, cytoplasmic Ca2+ buffering and sarcolemmal Ca2+ fluxes. Such analysis is important for understanding the cellular basis of inotropic interventions in cardiac muscle.
Figures




References
-
- Adams WA, Trafford AW, Eisner DA. 2,3-Butanedione monoxime (BDM) decreases sarcoplasmic reticulum Ca content by stimulating Ca release in isolated rat ventricular myocytes. Pflügers Archiv. 1998;436:776–781. - PubMed
-
- Bassani JWM, Yuan W, Bers DM. Fractional SR Ca release is regulated by trigger Ca and SR Ca content in cardiac myocytes. American Journal of Physiology. 1995;268:C1313–1329. - PubMed
-
- Bers DM, Berlin JR. Kinetics of [Ca2+]i decline in cardiac myocytes depend on peak [Ca2+]i. American Journal of Physiology. 1995;268:C271–277. - PubMed
-
- Bers DM, Bridge JH, Macleod KT. The mechanism of ryanodine action in rabbit ventricular muscle evaluated with Ca-selective microelectrodes and rapid cooling contractures. Canadian Journal of Physiology and Pharmacology. 1987;65:610–618. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous