

Targeted disruption of hormone-sensitive lipase results in male sterility and adipocyte hypertrophy, but not in obesity
- PMID: 10639158
- PMCID: PMC15409
- DOI: 10.1073/pnas.97.2.787
Targeted disruption of hormone-sensitive lipase results in male sterility and adipocyte hypertrophy, but not in obesity
Abstract
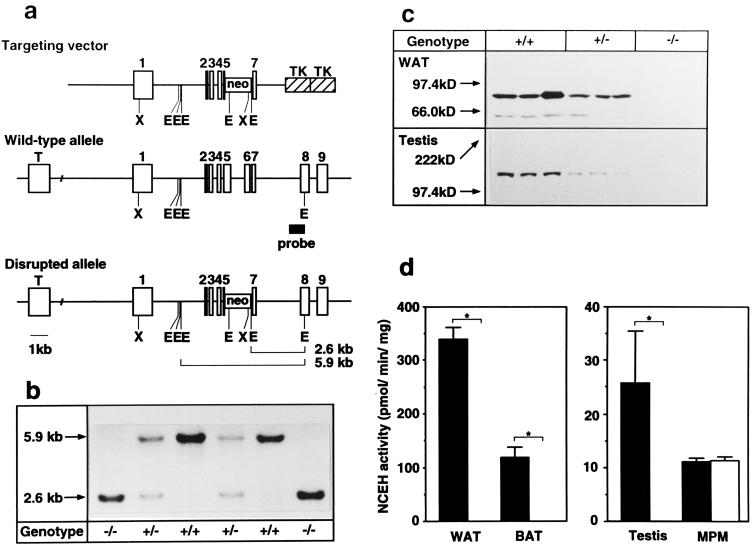
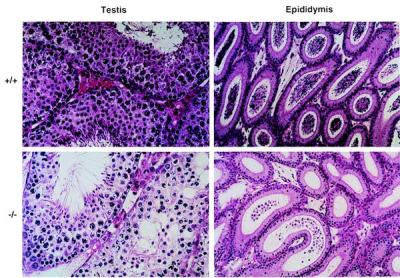
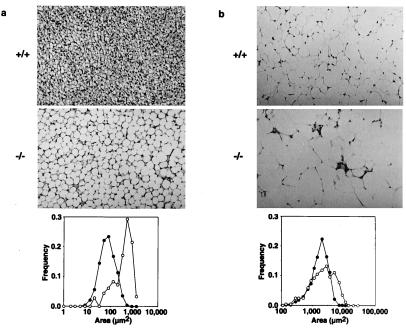
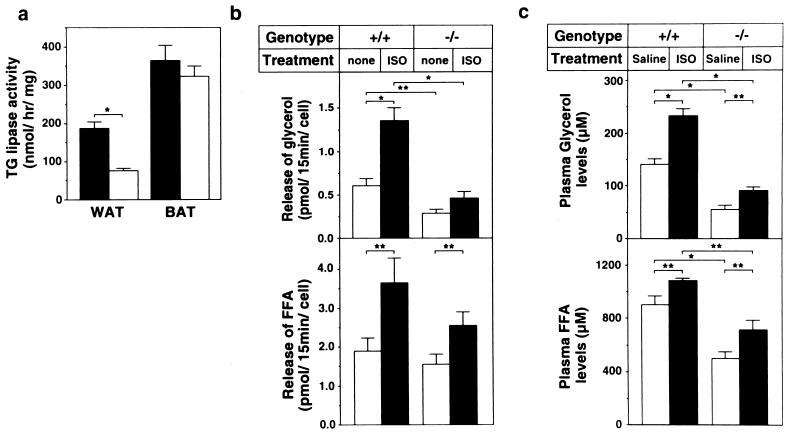
Hormone-sensitive lipase (HSL) is known to mediate the hydrolysis not only of triacylglycerol stored in adipose tissue but also of cholesterol esters in the adrenals, ovaries, testes, and macrophages. To elucidate its precise role in the development of obesity and steroidogenesis, we generated HSL knockout mice by homologous recombination in embryonic stem cells. Mice homozygous for the mutant HSL allele (HSL-/-) were superficially normal except that the males were sterile because of oligospermia. HSL-/- mice did not have hypogonadism or adrenal insufficiency. Instead, the testes completely lacked neutral cholesterol ester hydrolase (NCEH) activities and contained increased amounts of cholesterol ester. Many epithelial cells in the seminiferous tubules were vacuolated. NCEH activities were completely absent from both brown adipose tissue (BAT) and white adipose tissue (WAT) in HSL-/- mice. Consistently, adipocytes were significantly enlarged in the BAT (5-fold) and, to a lesser extent in the WAT (2-fold), supporting the concept that the hydrolysis of triacylglycerol was, at least in part, impaired in HSL-/- mice. The BAT mass was increased by 1.65-fold, but the WAT mass remained unchanged. Discrepancy of the size differences between cell and tissue suggests the heterogeneity of adipocytes. Despite these morphological changes, HSL-/- mice were neither obese nor cold sensitive. Furthermore, WAT from HSL-/- mice retained 40% of triacylglycerol lipase activities compared with the wild-type WAT. In conclusion, HSL is required for spermatogenesis but is not the only enzyme that mediates the hydrolysis of triacylglycerol stored in adipocytes.
Figures
Comment in
-
Another hormone-sensitive triglyceride lipase in fat cells?Proc Natl Acad Sci U S A. 2000 Jan 18;97(2):535-7. doi: 10.1073/pnas.97.2.535. Proc Natl Acad Sci U S A. 2000. PMID: 10639113 Free PMC article. No abstract available.
References
-
- Strålfors P, Olsson H, Belfrage P. In: The Enzymes. Boyer P D, Krebs E G, editors. London: Academic; 1987. pp. 147–177.
-
- Yeaman S J. Biochim Biophys Acta. 1990;1052:128–132. - PubMed
-
- Holm C, Kirchgessner T G, Svenson K L, Fredrikson G, Nilsson S, Miller C G, Shively J E, Heinzmann C, Sparkes R S, Mohandas T, et al. Science. 1988;241:1503–1506. - PubMed
-
- Fredrikson G, Strålfors P, Nilsson N, Belfrage P. J Biol Chem. 1981;256:6311–6320. - PubMed
-
- Eriksson H, Ridderstrale M, Degerman E, Ekholm D, Smith C J, Manganiello V C, Belfrage P, Tornqvist H. Biochim Biophys Acta. 1996;1266:101–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
