

Modulation of nucleotide sensitivity of ATP-sensitive potassium channels by phosphatidylinositol-4-phosphate 5-kinase
- PMID: 10639183
- PMCID: PMC15434
- DOI: 10.1073/pnas.97.2.937
Modulation of nucleotide sensitivity of ATP-sensitive potassium channels by phosphatidylinositol-4-phosphate 5-kinase
Abstract
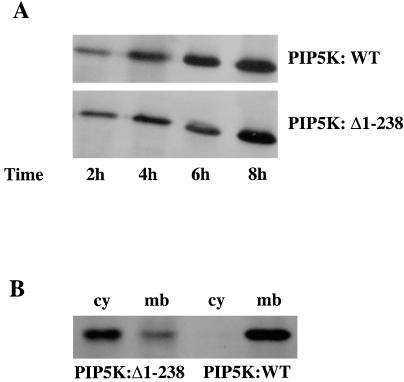
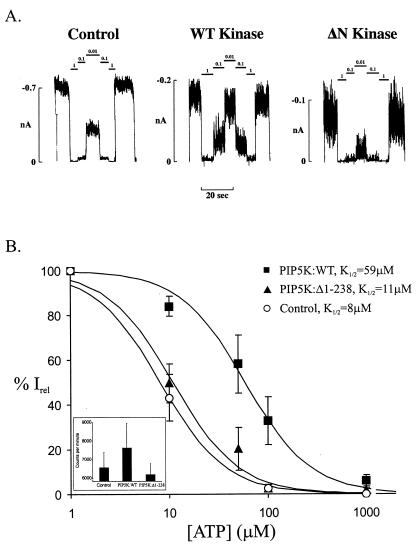
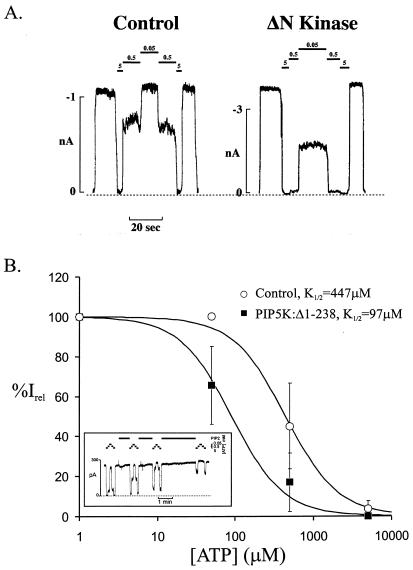
ATP-sensitive potassium channels (K(ATP) channels) regulate cell excitability in response to metabolic changes. K(ATP) channels are formed as a complex of a sulfonylurea receptor (SURx), a member of the ATP-binding cassette protein family, and an inward rectifier K(+) channel subunit (Kir6.x). Membrane phospholipids, in particular phosphatidylinositol (PI) 4,5-bisphosphate (PIP(2)), activate K(ATP) channels and antagonize ATP inhibition of K(ATP) channels when applied to inside-out membrane patches. To examine the physiological relevance of this regulatory mechanism, we manipulated membrane PIP(2) levels by expressing either the wild-type or an inactive form of PI-4-phosphate 5-kinase (PIP5K) in COSm6 cells and examined the ATP sensitivity of coexpressed K(ATP) channels. Channels from cells expressing the wild-type PIP5K have a 6-fold lower ATP sensitivity (K(1/2), the half maximal inhibitory concentration, approximately 60 microM) than the sensitivities from control cells (K(1/2) approximately 10 microM). An inactive form of the PIP5K had little effect on the K(1/2) of wild-type channels but increased the ATP-sensitivity of a mutant K(ATP) channel that has an intrinsically lower ATP sensitivity (from K(1/2) approximately 450 microM to K(1/2) approximately 100 microM), suggesting a decrease in membrane PIP(2) levels as a consequence of a dominant-negative effect of the inactive PIP5K. These results show that PIP5K activity, which regulates PIP(2) and PI-3,4,5-P(3) levels, is a significant determinant of the physiological nucleotide sensitivity of K(ATP) channels.
Figures
References
-
- Aguilar-Bryan L, Nichols C G, Wechsler S W, Clement J P, IV, Boyd A E, III, Gonzalez G, Herrera-Sosa H, Nguy K, Bryan J, Nelson D A. Science. 1995;268:423–426. - PubMed
-
- Inagaki N, Tsuura Y, Namba N, Masuda K, Gonoi T, Horie M, Seino Y, Mizuta M, Seino S. J Biol Chem. 1995;270:5691–5694. - PubMed
-
- Inagaki N, Gonoi T, Clement J P, IV, Namba N, Inazawa J, Gonzales G, Aguilar-Bryan L, Seino S, Bryan J. Science. 1995;270:1166–1170. - PubMed
-
- Clement J P, IV, Kunjilwar K, Gonzalez G, Schwanstecher M, Panten U, Aguilar-Bryan L, Bryan J. Neuron. 1997;18:827–838. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
