

HAESA, an Arabidopsis leucine-rich repeat receptor kinase, controls floral organ abscission
- PMID: 10640280
- PMCID: PMC316334
HAESA, an Arabidopsis leucine-rich repeat receptor kinase, controls floral organ abscission
Abstract
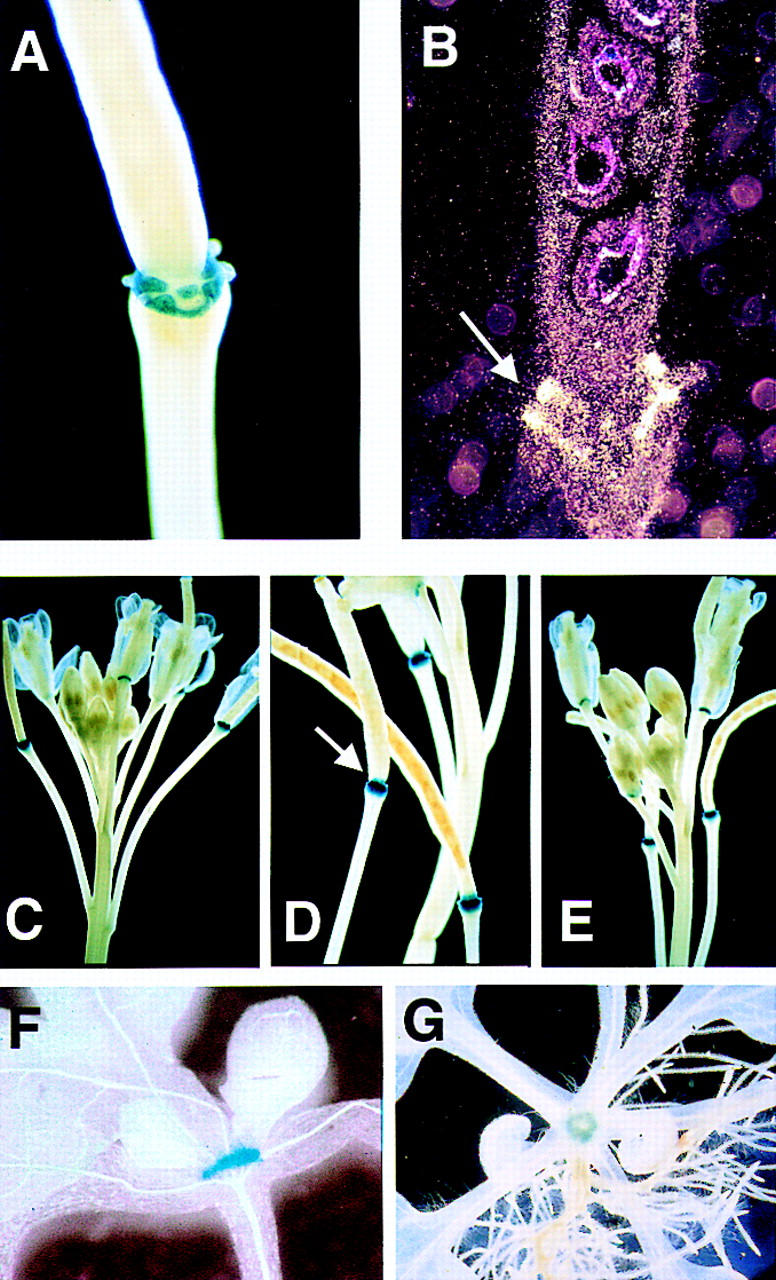
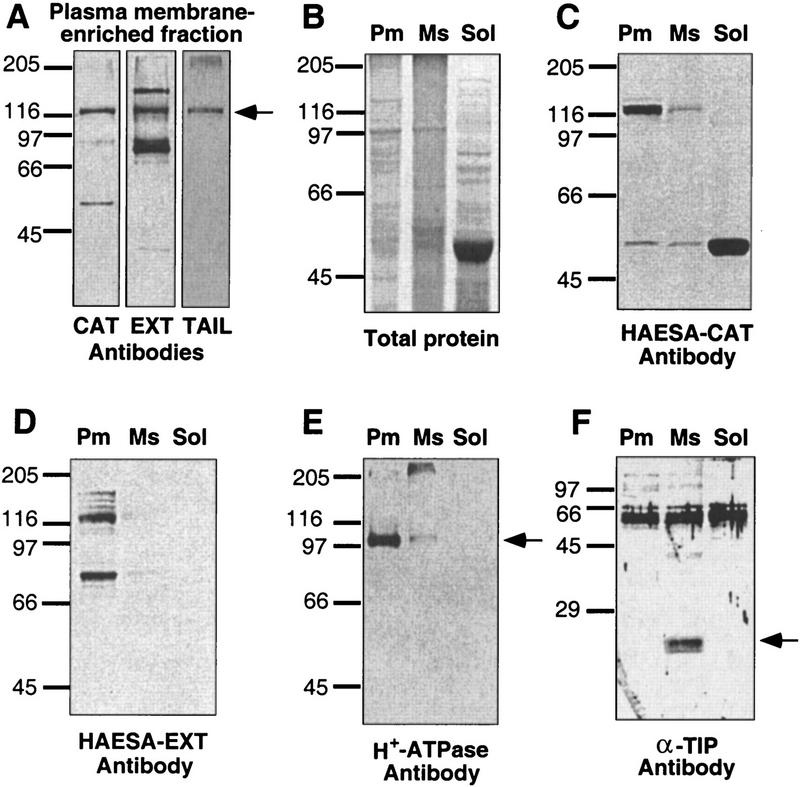
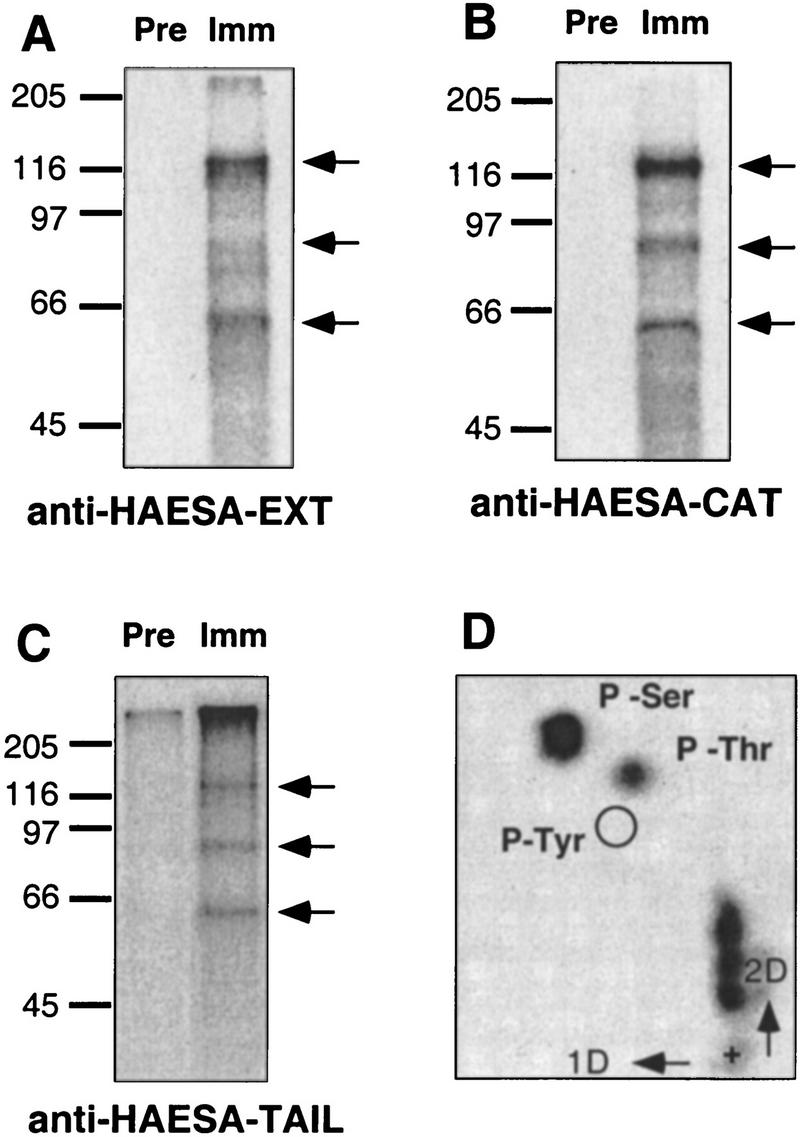
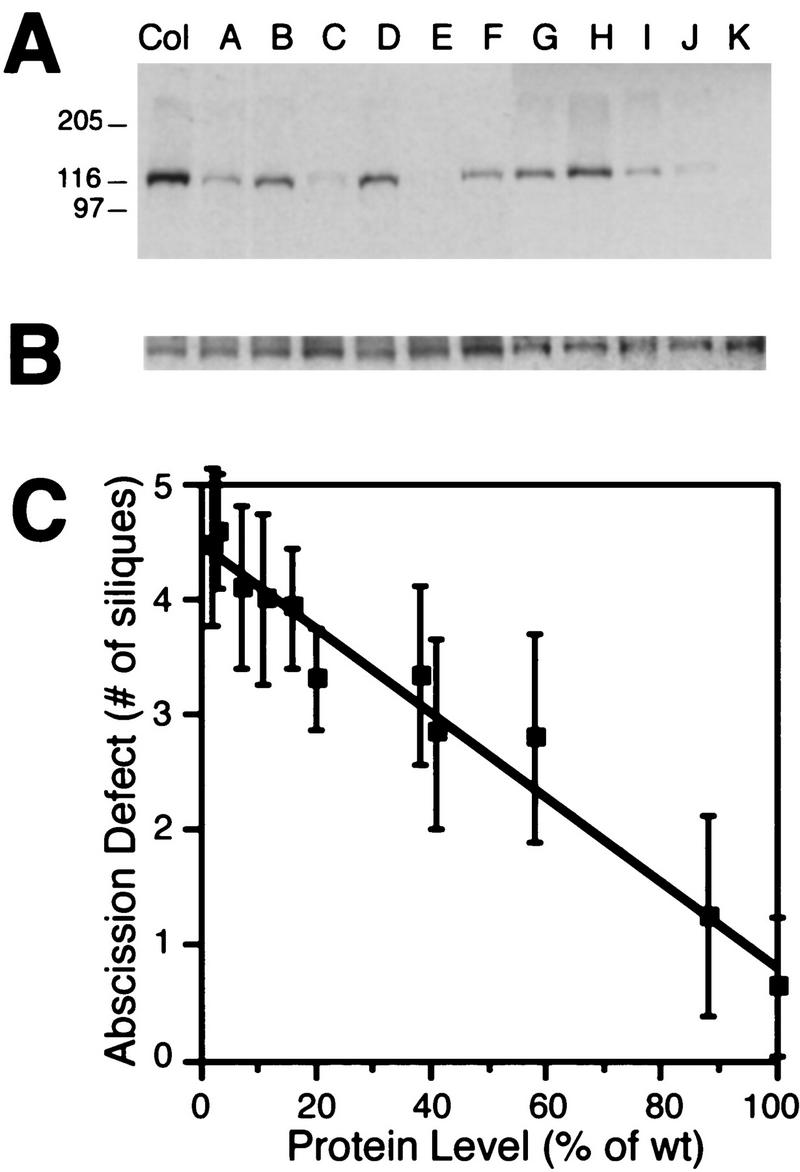
Abcission, the natural shedding of leaves, flowers and fruits, is a fundamental component of plant development. Abscission is a highly regulated process that occurs at distinct zones of cells that undergo enlargement and subsequent separation. Although some components of abscission, including accumulation of the hormone ethylene and cell wall-degrading enzymes, have been described, the regulatory pathways remain largely unknown. In this paper we describe a critical component required for floral organ abscission in Arabidopsis thaliana, the receptor-like protein kinase HAESA. Histochemical analysis of transgenic plants harboring a HAESA promoter:: beta-glucuronidase reporter gene and in situ RNA hybridization experiments show HAESA expression in the abscission zones where the sepals, petals, and stamens attach to the receptacle, at the base of pedicels, and at the base of petioles where leaves attach to the stem. Immunodetection, immunoprecipitation, and protein kinase activity assays reveal HAESA is a plasma membrane serine/threonine protein kinase. The reduction of function of HAESA in transgenic plants harboring an antisense construct results in delayed abscission of floral organs, and the severity of the phenotype is directly correlated with the level of HAESA protein. These results demonstrate that HAESA functions in developmentally regulated floral organ abscission.
Figures
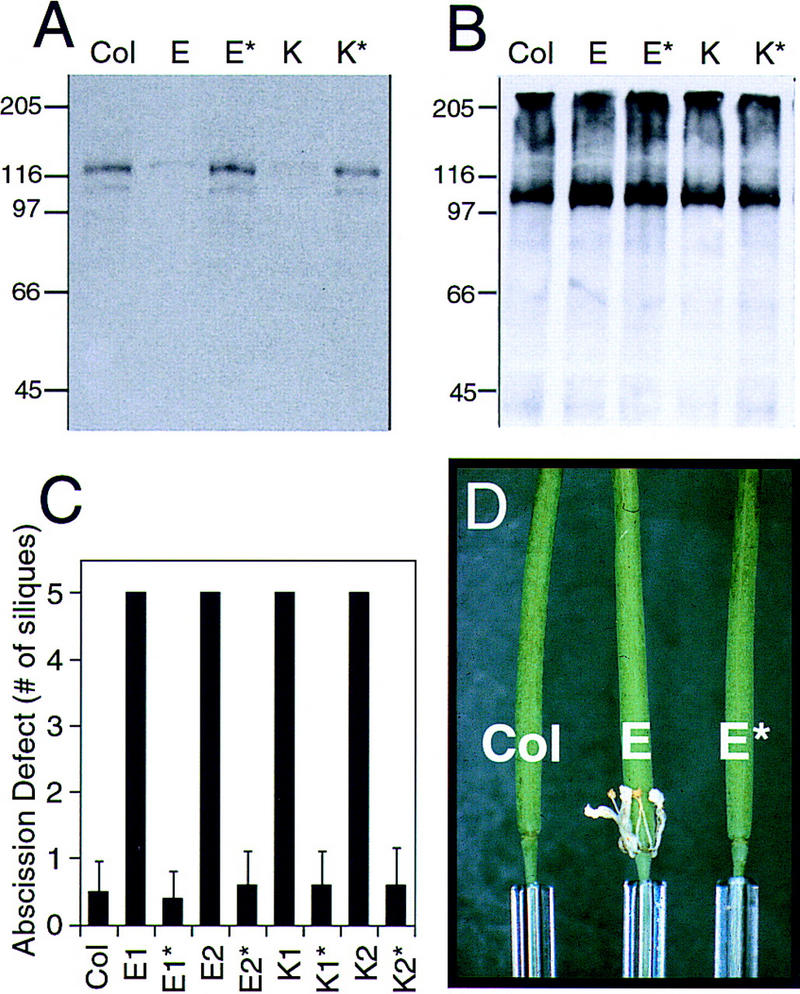
References
-
- Altmann T. Recent advances in brassinosteroid molecular genetics. Curr Opin Plant Biol. 1998;1:378–383. - PubMed
-
- An G, Ebert PR, Mitra A, Ha SB. Binary vectors. Plant Mol Biol Manual. 1988;A3:1–19.
-
- Bechtold N, Bouchez D. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. In: Potrykus I, Spangenberg G, editors. Gene transfer to plants. New York, NY: Springer Verlag; 1994. p. 361. - PubMed
-
- Becraft PW. Receptor kinases in plant development. Trends Plant Sci. 1998;3:384–388.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases