

Requirement of aquaporin-1 for NaCl-driven water transport across descending vasa recta
- PMID: 10642600
- PMCID: PMC382858
- DOI: 10.1172/JCI8214
Requirement of aquaporin-1 for NaCl-driven water transport across descending vasa recta
Abstract
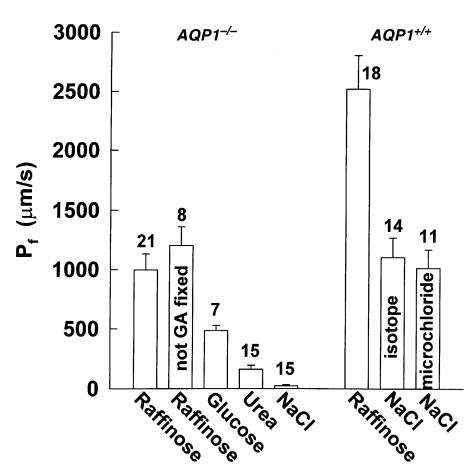
Deletion of AQP1 in mice results in diminished urinary concentrating ability, possibly related to reduced NaCl- and urea gradient-driven water transport across the outer medullary descending vasa recta (OMDVR). To quantify the role of AQP1 in OMDVR water transport, we measured osmotically driven water permeability in vitro in microperfused OMDVR from wild-type, AQP1 heterozygous, and AQP1 knockout mice. OMDVR diameters in AQP1(-/-) mice were 1.9-fold greater than in AQP1(+/+) mice. Osmotic water permeability (P(f)) in response to a 200 mM NaCl gradient (bath > lumen) was reduced about 2-fold in AQP1(+/-) mice and by more than 50-fold in AQP1(-/-) mice. P(f) increased from 1015 to 2527 microm/s in AQP1(+/+) mice and from 22 to 1104 microm/s in AQP1(-/-) mice when a raffinose rather than an NaCl gradient was used. This information, together with p-chloromercuribenzenesulfonate inhibition measurements, suggests that nearly all NaCl-driven water transport occurs by a transcellular route through AQP1, whereas raffinose-driven water transport also involves a parallel, AQP1-independent, mercurial-insensitive pathway. Interestingly, urea was also able to drive water movement across the AQP1-independent pathway. Diffusional permeabilities to small hydrophilic solutes were comparable in AQP1(+/+) and AQP1(-/-) mice but higher than those previously measured in rats. In a mathematical model of the medullary microcirculation, deletion of AQP1 resulted in diminished concentrating ability due to enhancement of medullary blood flow, partially accounting for the observed urine-concentrating defect.
Figures






References
-
- Pallone TL, Robertson CR, Jamison RL. Renal medullary microcirculation. Physiol Rev. 1990;70:885–920. - PubMed
-
- Nielsen S, et al. Aquaporin CHIP water channels in short and long loop descending thin limb and in descending vasa recta in rat kidney. Am J Physiol. 1995;268:F1023–F1037. - PubMed
-
- Pallone TL, Nielsen S, Silldorff EP, Yang S. Diffusive transport of solute in the rat medullary microcirculation. Am J Physiol. 1995;269:F55–F63. - PubMed
-
- Pallone TL. Characterization of the urea transporter in outer medullary descending vasa recta. Am J Physiol. 1994;267:R260–R267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
