

Hepatitis C virus core protein interacts with 14-3-3 protein and activates the kinase Raf-1
- PMID: 10644344
- PMCID: PMC111649
- DOI: 10.1128/jvi.74.4.1736-1741.2000
Hepatitis C virus core protein interacts with 14-3-3 protein and activates the kinase Raf-1
Abstract
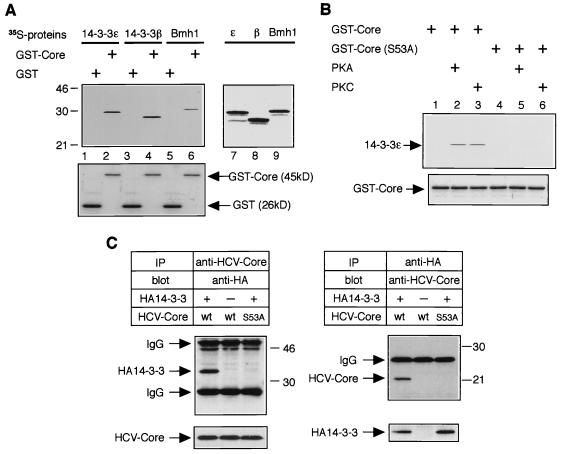
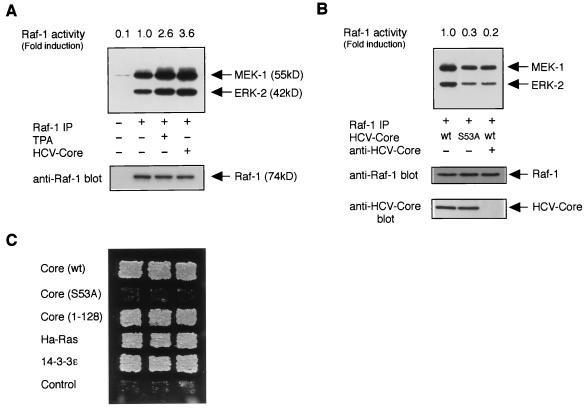
Persistent hepatitis C virus (HCV) infection is a major cause of chronic liver dysfunction in humans and is epidemiologically closely associated with the development of human hepatocellular carcinoma. Among HCV components, core protein has been reported to be implicated in cell growth regulation both in vitro and in vivo, although mechanisms explaining those effects are still unclear. In the present study, we identified that members of the 14-3-3 protein family associate with HCV core protein. 14-3-3 protein bound to HCV core protein in a phosphoserine-dependent manner. Introduction of HCV core protein caused a substantial increase in Raf-1 kinase activity in HepG2 cells and in a yeast genetic assay. Furthermore, the HCV core-14-3-3 interaction was essential for Raf-1 kinase activation by HCV core protein. These results suggest that HCV core protein may represent a novel type of Raf-1 kinase-activating protein through its interaction with 14-3-3 protein and may contribute to hepatocyte growth regulation.
Figures

References
-
- Aitken A. 14-3-3 proteins on the MAP. Trends Cell Biol. 1996;6:341–347. - PubMed
-
- Alter H J, Purcell R H, Shih J W, Melpolder J C, Houghton M, Choo Q-L, Kuo G. Detection of antibody to hepatitis C virus in prospectively followed transfusion recipients with acute and chronic non-A, non-B hepatitis. N Engl J Med. 1989;321:1494–1500. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
