

Anterograde transport of herpes simplex virus type 1 in cultured, dissociated human and rat dorsal root ganglion neurons
- PMID: 10644356
- PMCID: PMC111661
- DOI: 10.1128/jvi.74.4.1827-1839.2000
Anterograde transport of herpes simplex virus type 1 in cultured, dissociated human and rat dorsal root ganglion neurons
Abstract
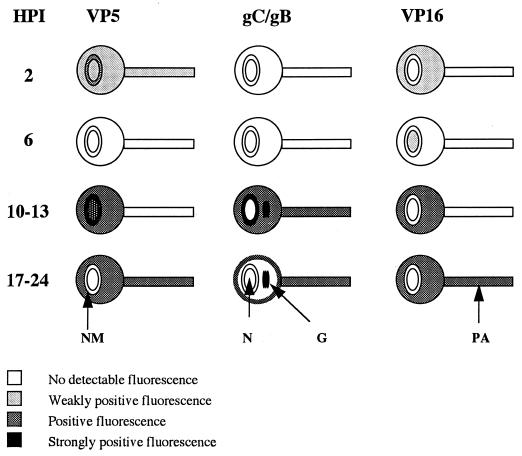
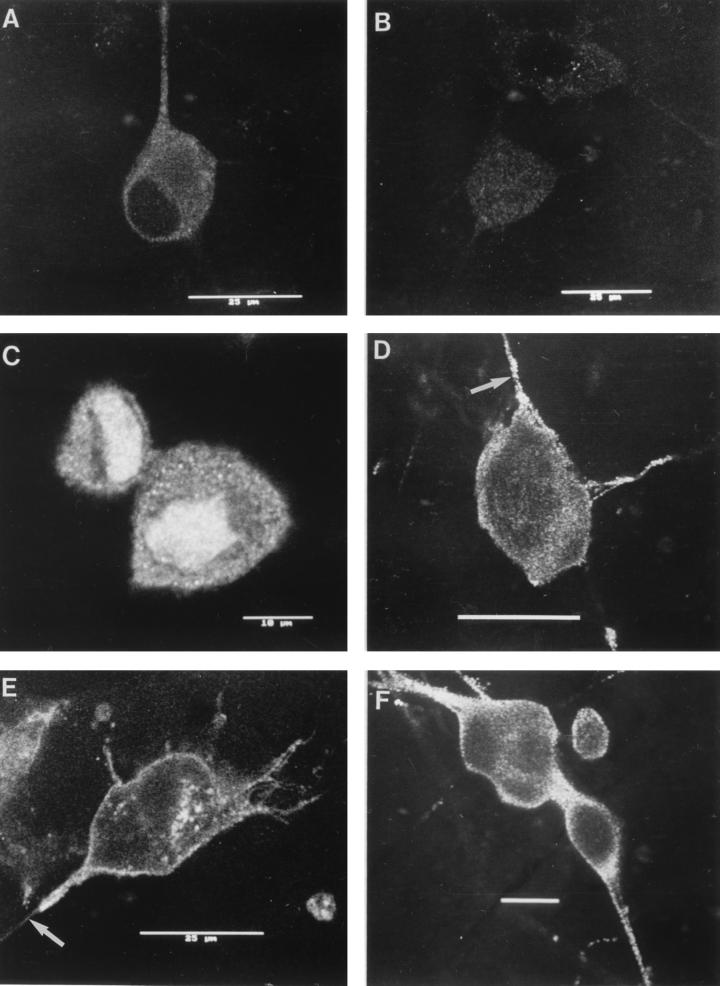
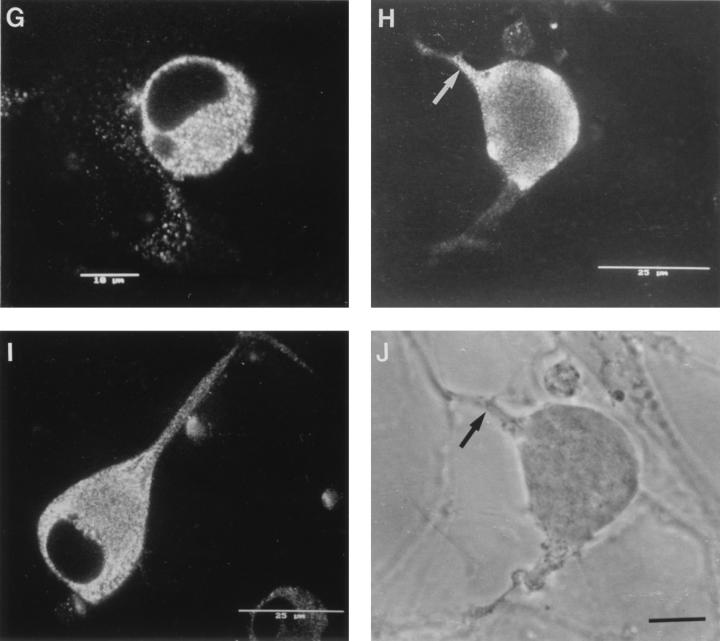
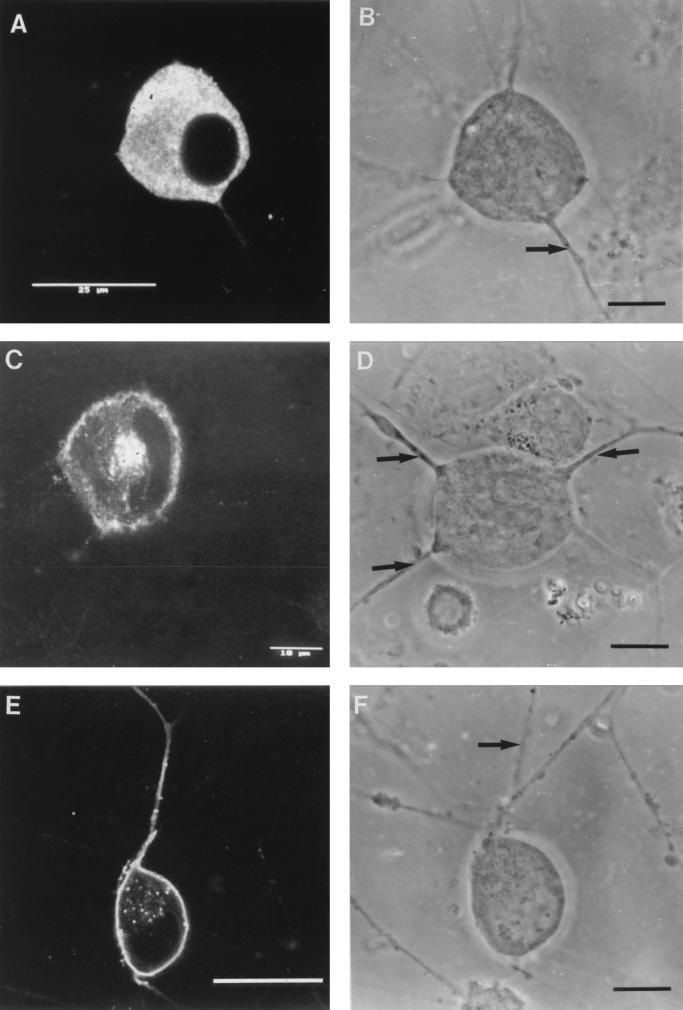
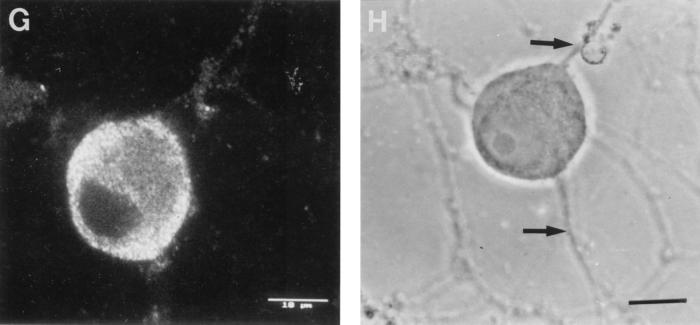
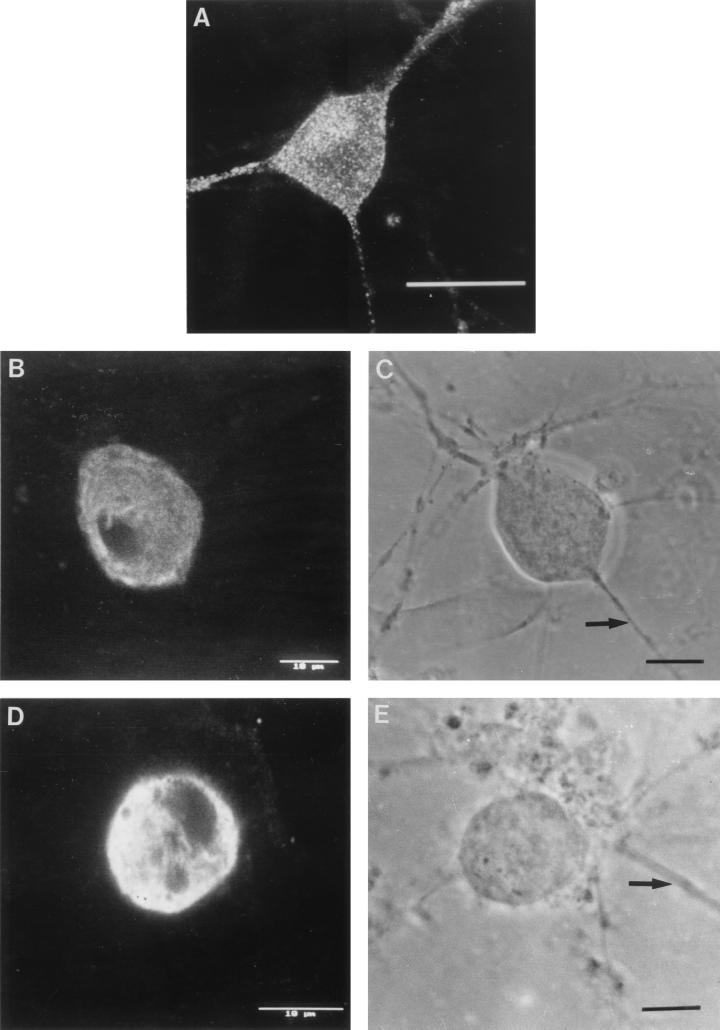
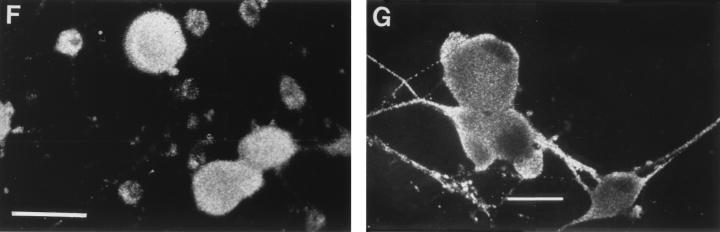
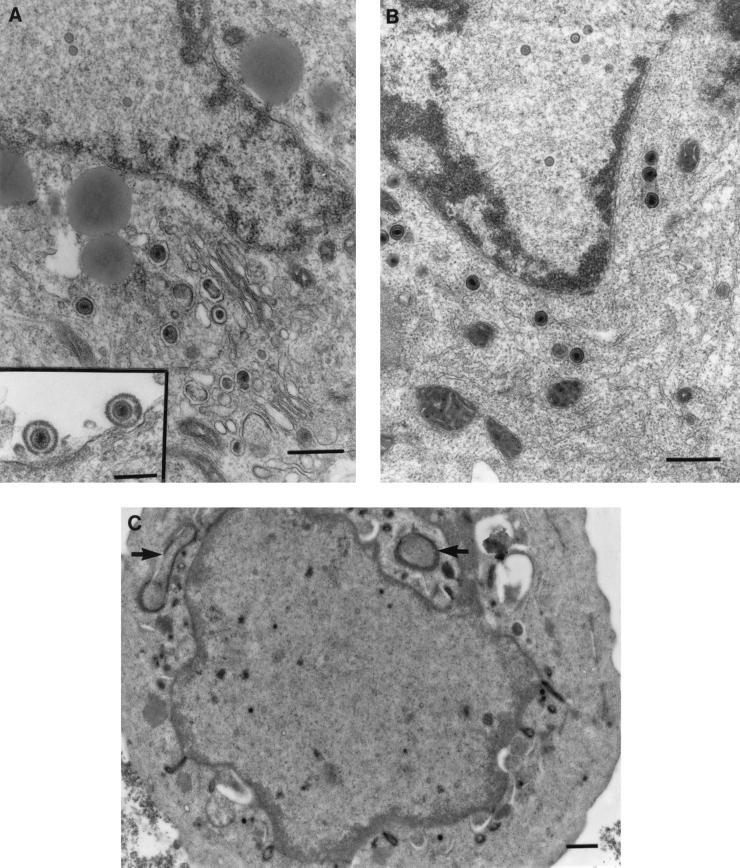
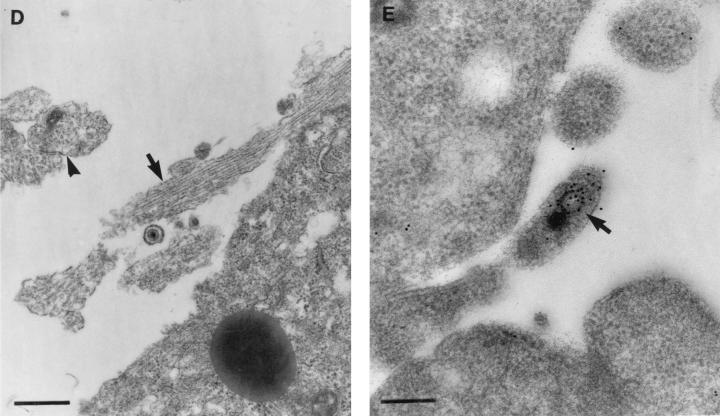
The mechanism of anterograde transport of herpes simplex virus was studied in cultured dissociated human and rat dorsal root ganglion neurons. The neurons were infected with HSV-1 to examine the distribution of capsid (VP5), tegument (VP16), and glycoproteins (gC and gB) at 2, 6, 10, 13, 17, and 24 h postinfection (p.i.) with or without nocodazole (a microtubule depolymerizer) or brefeldin A (a Golgi inhibitor). Retrogradely transported VP5 was detected in the cytoplasm of the cell body up to the nuclear membrane at 2 h p.i. It was first detected de novo in the nucleus and cytoplasm at 10 h p.i., the axon hillock at 13 h p.i., and the axon at 15 to 17 h p.i. gC and gB were first detected de novo in the cytoplasm and the axon hillock at 10 h p.i. and then in the axon at 13 h p.i., which was always earlier than the detection of VP5. De novo-synthesized VP16 was first detected in the cytoplasm at 10 to 13 h p.i. and in the axon at 16 to 17 h p.i. Nocodazole inhibited the transport of all antigens, VP5, VP16, and gC or gB. The kinetics of inhibition of VP5 and gC could be dissociated. Brefeldin A inhibited the transport of gC or gB and VP16 but not VP5 into axons. Transmission immunoelectron microscopy confirmed that there were unenveloped nucleocapsids in the axon with or without brefeldin A. These findings demonstrate that glycoproteins and capsids, associated with tegument proteins, are transported by different pathways with slightly differing kinetics from the nucleus to the axon. Furthermore, axonal anterograde transport of the nucleocapsid can proceed despite the loss of most VP16.
Figures
Similar articles
-
In rat dorsal root ganglion neurons, herpes simplex virus type 1 tegument forms in the cytoplasm of the cell body.J Virol. 2002 Oct;76(19):9934-51. doi: 10.1128/jvi.76.19.9934-9951.2002. J Virol. 2002. PMID: 12208970 Free PMC article.
-
Anterograde transport of herpes simplex virus proteins in axons of peripheral human fetal neurons: an immunoelectron microscopy study.J Virol. 1999 Oct;73(10):8503-11. doi: 10.1128/JVI.73.10.8503-8511.1999. J Virol. 1999. PMID: 10482603 Free PMC article.
-
Herpes simplex virus type 1 accumulation, envelopment, and exit in growth cones and varicosities in mid-distal regions of axons.J Virol. 2006 Apr;80(7):3592-606. doi: 10.1128/JVI.80.7.3592-3606.2006. J Virol. 2006. PMID: 16537627 Free PMC article.
-
Transport and egress of herpes simplex virus in neurons.Rev Med Virol. 2008 Jan-Feb;18(1):35-51. doi: 10.1002/rmv.560. Rev Med Virol. 2008. PMID: 17992661 Review.
-
Anterograde transport of α-herpesviruses in neuronal axons.Virology. 2021 Jul;559:65-73. doi: 10.1016/j.virol.2021.02.011. Epub 2021 Mar 4. Virology. 2021. PMID: 33836340 Review.
Cited by
-
Herpes Simplex Virus Type 1 Interactions with the Interferon System.Int J Mol Sci. 2020 Jul 21;21(14):5150. doi: 10.3390/ijms21145150. Int J Mol Sci. 2020. PMID: 32708188 Free PMC article. Review.
-
Targeted Promoter Replacement Reveals That Herpes Simplex Virus Type-1 and 2 Specific VP16 Promoters Direct Distinct Rates of Entry Into the Lytic Program in Sensory Neurons in vivo.Front Microbiol. 2019 Jul 24;10:1624. doi: 10.3389/fmicb.2019.01624. eCollection 2019. Front Microbiol. 2019. PMID: 31396171 Free PMC article.
-
A hitchhiker's guide to the nervous system: the complex journey of viruses and toxins.Nat Rev Microbiol. 2010 Sep;8(9):645-55. doi: 10.1038/nrmicro2395. Nat Rev Microbiol. 2010. PMID: 20706281 Review.
-
Alpha and gamma interferons inhibit herpes simplex virus type 1 infection and spread in epidermal cells after axonal transmission.J Virol. 2001 Dec;75(23):11821-6. doi: 10.1128/JVI.75.23.11821-11826.2001. J Virol. 2001. PMID: 11689663 Free PMC article.
-
Reconstitution of herpes simplex virus microtubule-dependent trafficking in vitro.J Virol. 2006 May;80(9):4264-75. doi: 10.1128/JVI.80.9.4264-4275.2006. J Virol. 2006. PMID: 16611885 Free PMC article.
References
-
- Chatterjee S, Sarkar S. Studies on endoplasmic reticulum-Golgi complex cycling pathway in herpes simplex virus-infected and brefeldin A-treated human fibroblast cells. Virology. 1992;191:327–337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
