

Characterization of gammaherpesvirus 68 gene 50 transcription
- PMID: 10644377
- PMCID: PMC111682
- DOI: 10.1128/jvi.74.4.2029-2037.2000
Characterization of gammaherpesvirus 68 gene 50 transcription
Abstract
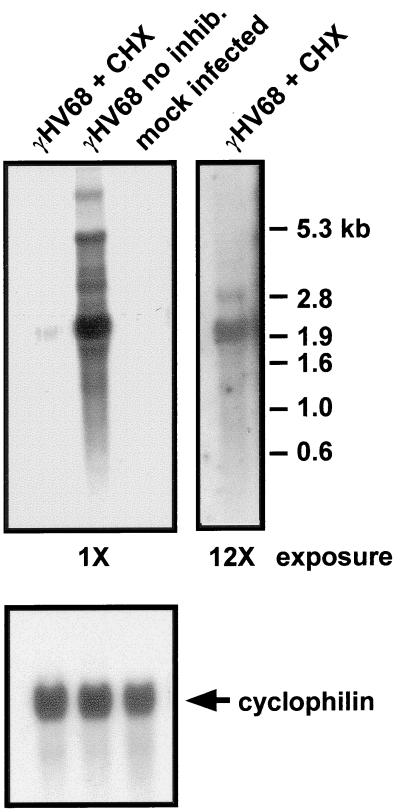
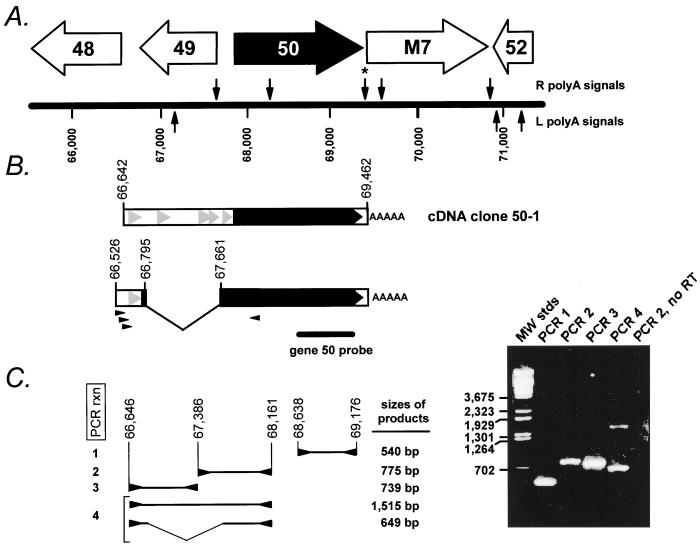
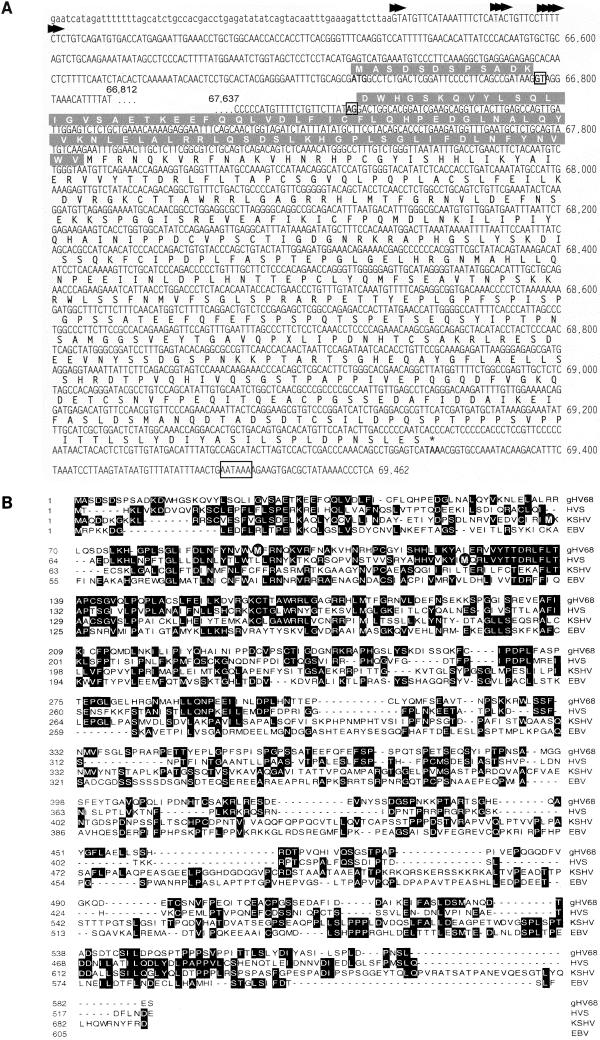
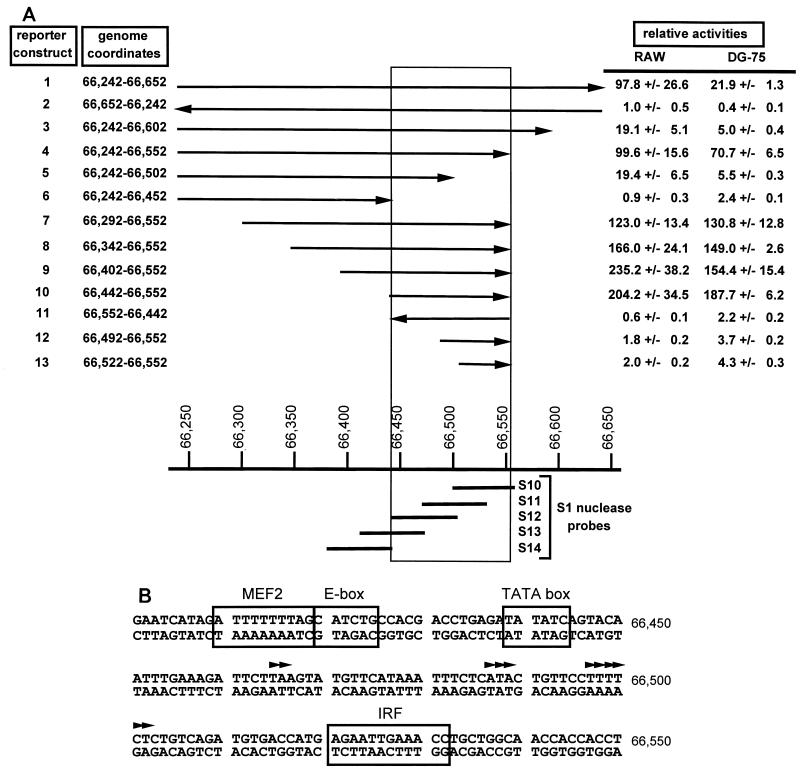
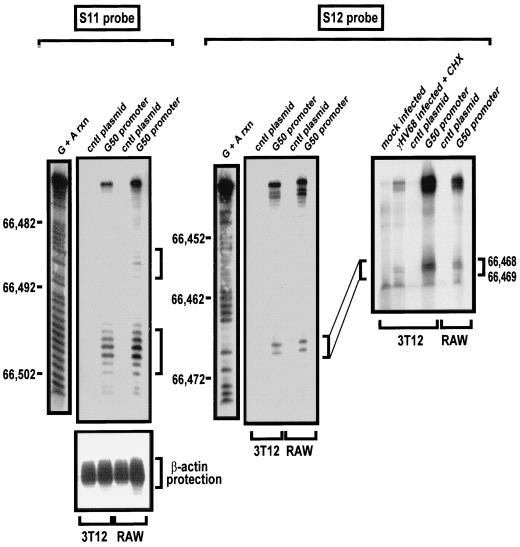
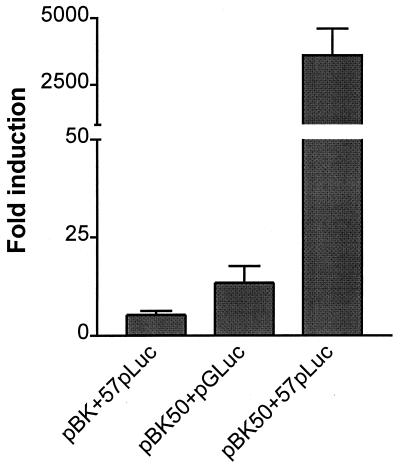
Gene 50 is the only immediate-early gene that appears to be conserved among the characterized gammaherpesviruses. It has recently been demonstrated for the human viruses Epstein-Barr virus (EBV) and Kaposi's sarcoma-associated herpesvirus (KSHV) that ectopic expression of the gene 50-encoded product in some latently infected cell lines can lead to the induction of virus replication, indicating that gene 50 is likely to play a pivotal role in regulating gammaherpesvirus reactivation. Here we demonstrate that the murine gammaherpesvirus 68 (gammaHV68) gene 50 is an immediate-early gene and that transcription of gammaHV68 gene 50 leads to the production of both spliced and unspliced forms of the gene 50 transcript. Splicing of the transcript near the 5' end serves to extend the gene 50 open reading frame, as has been observed for the gene 50 transcripts encoded by KSHV and herpesvirus saimiri (Whitehouse et al., J. Virol. 71:2550-2554, 1997; Lukac et al., Virology 252:304-312, 1998; Sun et al., Proc. Natl. Acad. Sci. USA 95:10866-10871, 1998). Reverse transcription-PCR analyses, coupled with S1 nuclease protection assays, provided evidence that gene 50 transcripts initiate at several sites within the region from bp 66468 to 66502 in the gammaHV68 genome. Functional characterization of the region upstream of the putative gene 50 transcription initiation site demonstrated orientation-dependent promoter activity and identified a 110-bp region (bp 66442 to 66552) encoding the putative gene 50 promoter. Finally, we demonstrate that the gammaHV68 gene 50 can transactivate the gammaHV68 gene 57 promoter, a known early gene target of the gene 50-encoded transactivator in other gammaherpesviruses. These studies show that the gammaHV68 gene 50 shares several important molecular similarities with the gene 50 homologs in other gammaherpesviruses and thus provides an impetus for future studies analyzing the role of the gammaHV68 gene 50-encoded protein in acute virus replication and reactivation from latency in vivo.
Figures
Similar articles
-
Disruption of gammaherpesvirus 68 gene 50 demonstrates that Rta is essential for virus replication.J Virol. 2003 May;77(10):5731-9. doi: 10.1128/jvi.77.10.5731-5739.2003. J Virol. 2003. PMID: 12719566 Free PMC article.
-
Rta of murine gammaherpesvirus 68 reactivates the complete lytic cycle from latency.J Virol. 2000 Apr;74(8):3659-67. doi: 10.1128/jvi.74.8.3659-3667.2000. J Virol. 2000. PMID: 10729142 Free PMC article.
-
Identification of Novel Kaposi's Sarcoma-Associated Herpesvirus Orf50 Transcripts: Discovery of New RTA Isoforms with Variable Transactivation Potential.J Virol. 2016 Dec 16;91(1):e01434-16. doi: 10.1128/JVI.01434-16. Print 2017 Jan 1. J Virol. 2016. PMID: 27795414 Free PMC article.
-
The Rta/Orf50 transactivator proteins of the gamma-herpesviridae.Curr Top Microbiol Immunol. 2007;312:71-100. doi: 10.1007/978-3-540-34344-8_3. Curr Top Microbiol Immunol. 2007. PMID: 17089794 Review.
-
Unraveling immunity to gamma-herpesviruses: a new model for understanding the role of immunity in chronic virus infection.Curr Opin Immunol. 1999 Aug;11(4):371-9. doi: 10.1016/s0952-7915(99)80063-6. Curr Opin Immunol. 1999. PMID: 10448140 Review.
Cited by
-
Gammaherpesvirus lytic gene expression as characterized by DNA array.J Virol. 2002 Jun;76(12):6244-56. doi: 10.1128/jvi.76.12.6244-6256.2002. J Virol. 2002. PMID: 12021358 Free PMC article.
-
Delta-9 tetrahydrocannabinol (THC) inhibits lytic replication of gamma oncogenic herpesviruses in vitro.BMC Med. 2004 Sep 15;2:34. doi: 10.1186/1741-7015-2-34. BMC Med. 2004. PMID: 15369590 Free PMC article.
-
Disruption of gammaherpesvirus 68 gene 50 demonstrates that Rta is essential for virus replication.J Virol. 2003 May;77(10):5731-9. doi: 10.1128/jvi.77.10.5731-5739.2003. J Virol. 2003. PMID: 12719566 Free PMC article.
-
Murine gammaherpesvirus 68 open reading frame 24 is required for late gene expression after DNA replication.J Virol. 2007 Jun;81(12):6761-4. doi: 10.1128/JVI.02726-06. Epub 2007 Mar 28. J Virol. 2007. PMID: 17392360 Free PMC article.
-
Murine gammaherpesvirus 68 has evolved gamma interferon and stat1-repressible promoters for the lytic switch gene 50.J Virol. 2010 Apr;84(7):3711-7. doi: 10.1128/JVI.02099-09. Epub 2010 Jan 13. J Virol. 2010. PMID: 20071569 Free PMC article.
References
-
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. - PubMed
-
- Daibata M, Speck S H, Mulder C, Sairenji T. Regulation of the BZLF1 promoter of Epstein-Barr virus by second messengers in anti-immunoglobulin-treated B cells. Virology. 1994;198:446–454. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
