

Insertional inactivation of genes encoding components of the sodium-type flagellar motor and switch of Vibrio parahaemolyticus
- PMID: 10648530
- PMCID: PMC94380
- DOI: 10.1128/JB.182.4.1035-1045.2000
Insertional inactivation of genes encoding components of the sodium-type flagellar motor and switch of Vibrio parahaemolyticus
Abstract
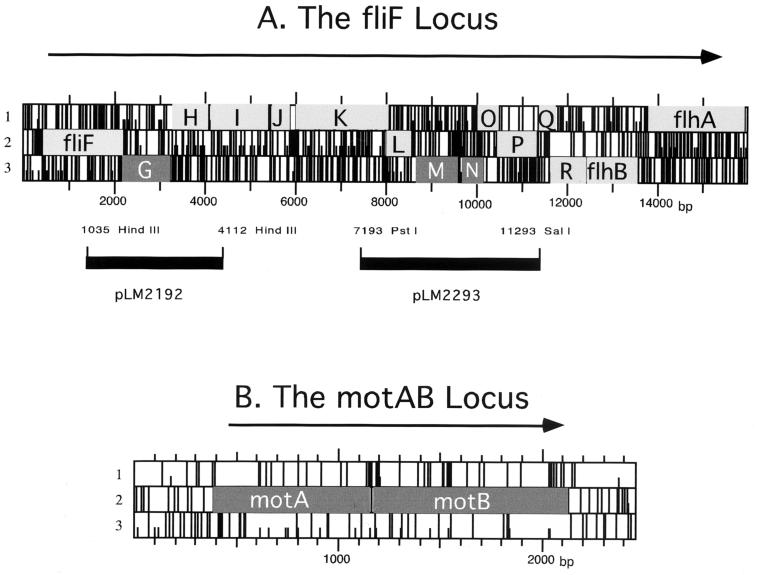
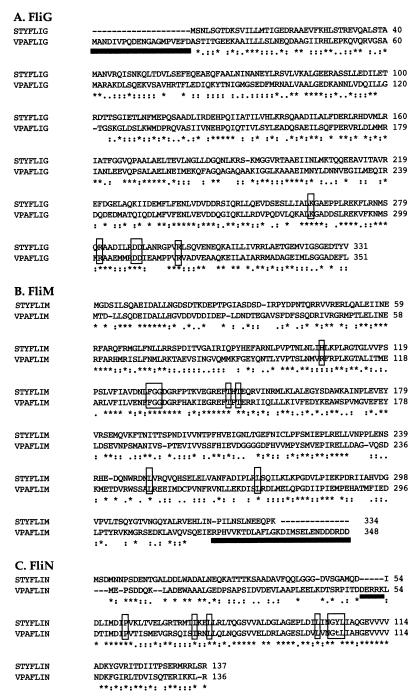
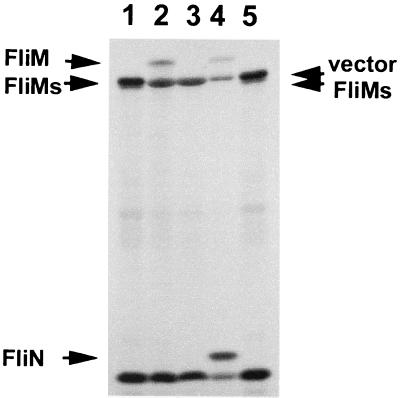
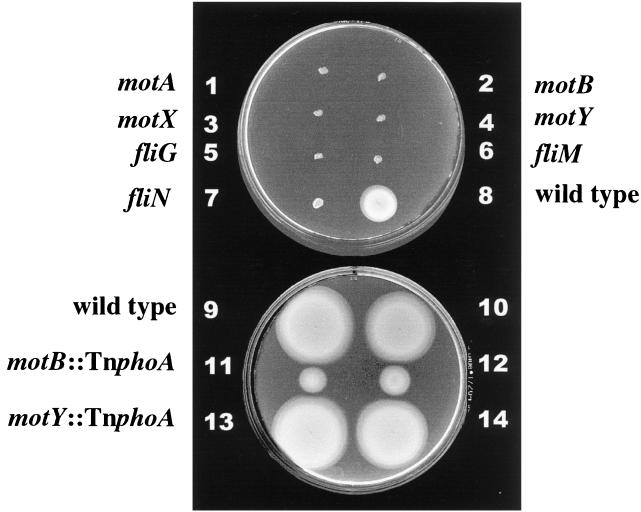
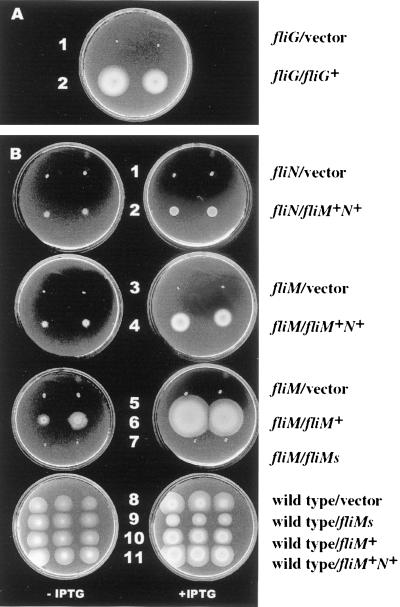
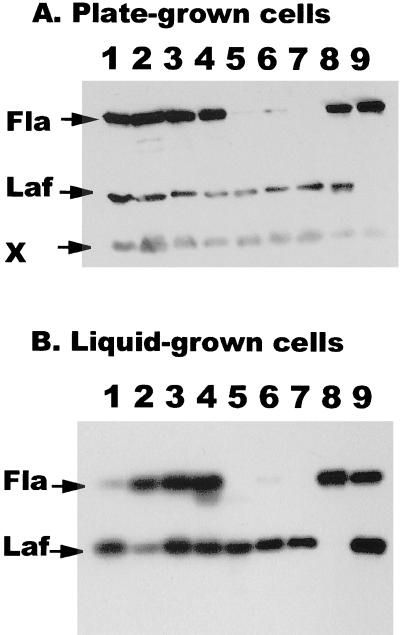
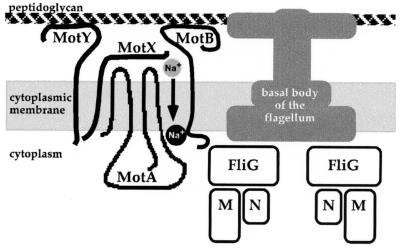
Vibrio parahaemolyticus possesses two types of flagella, polar and lateral, powered by distinct energy sources, which are derived from the sodium and proton motive forces, respectively. Although proton-powered flagella in Escherichia coli and Salmonella enterica serovar Typhimurium have been extensively studied, the mechanism of torque generation is still not understood. Molecular knowledge of the structure of the sodium-driven motor is only now being developed. In this work, we identify the switch components, FliG, FliM, and FliN, of the sodium-type motor. This brings the total number of genes identified as pertinent to polar motor function to seven. Both FliM and FliN possess charged domains not found in proton-type homologs; however, they can interact with the proton-type motor of E. coli to a limited extent. Residues known to be critical for torque generation in the proton-type motor are conserved in the sodium-type motor, suggesting a common mechanism for energy transfer at the rotor-stator interface regardless of the driving force powering rotation. Mutants representing a complete panel of insertionally inactivated switch and motor genes were constructed. All of these mutants were defective in sodium-driven swimming motility. Alkaline phosphatase could be fused to the C termini of MotB and MotY without abolishing motility, whereas deletion of the unusual, highly charged C-terminal domain of FliM disrupted motor function. All of the mutants retained proton-driven, lateral motility over surfaces. Thus, although central chemotaxis genes are shared by the polar and lateral systems, genes encoding the switch components, as well as the motor genes, are distinct for each motility system.
Figures

References
-
- Atsumi T, McCarter L, Imae Y. Polar and lateral flagellar motors of marine Vibrio are driven by different ion membrane forces. Nature (London) 1992;355:182–184. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
