

Novel WD-repeat protein Mip1p facilitates function of the meiotic regulator Mei2p in fission yeast
- PMID: 10648609
- PMCID: PMC85253
- DOI: 10.1128/MCB.20.4.1234-1242.2000
Novel WD-repeat protein Mip1p facilitates function of the meiotic regulator Mei2p in fission yeast
Abstract
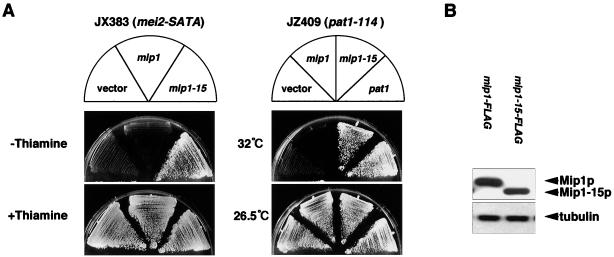
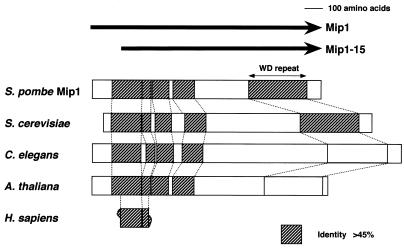
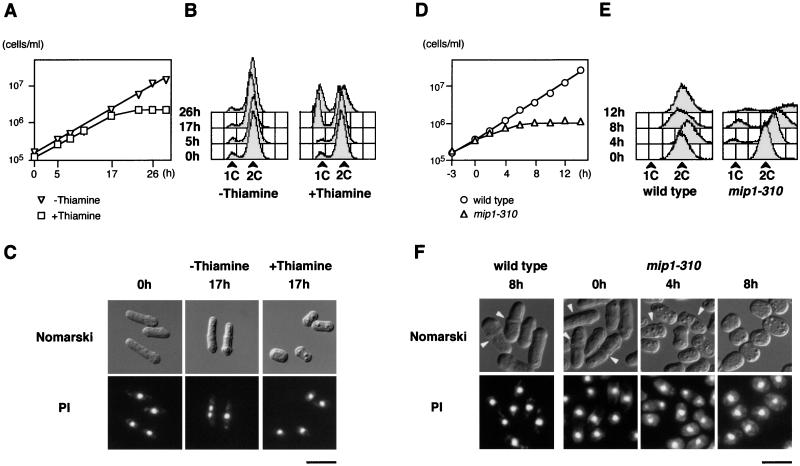
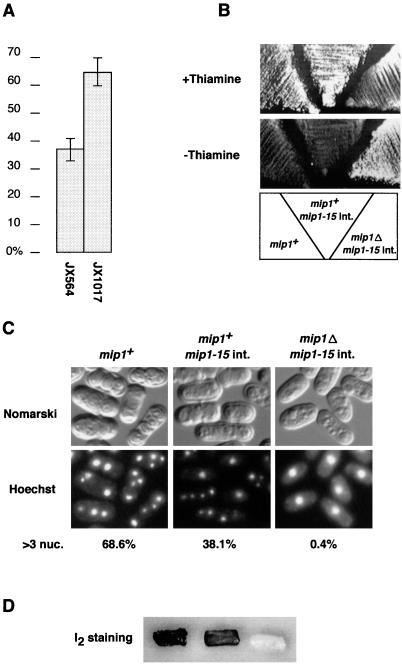
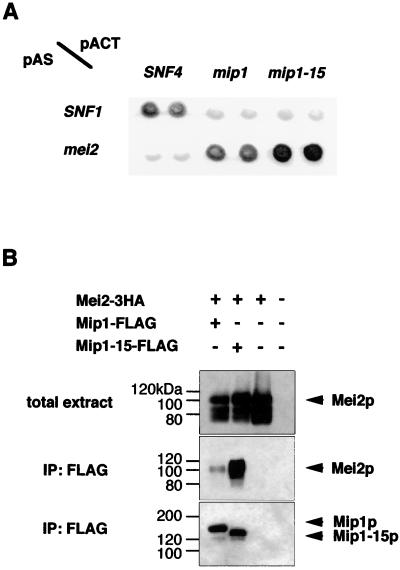
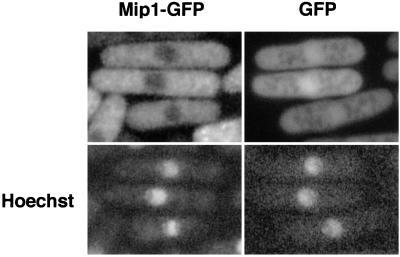
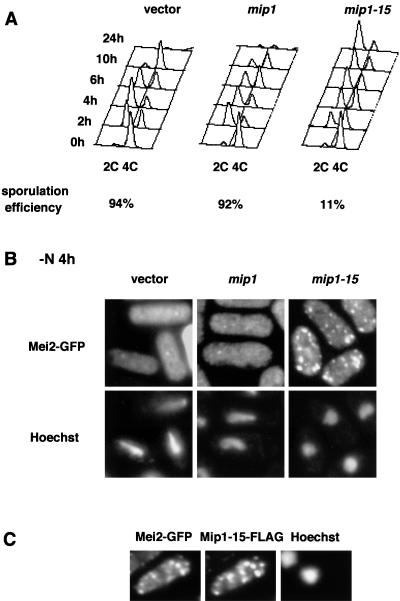
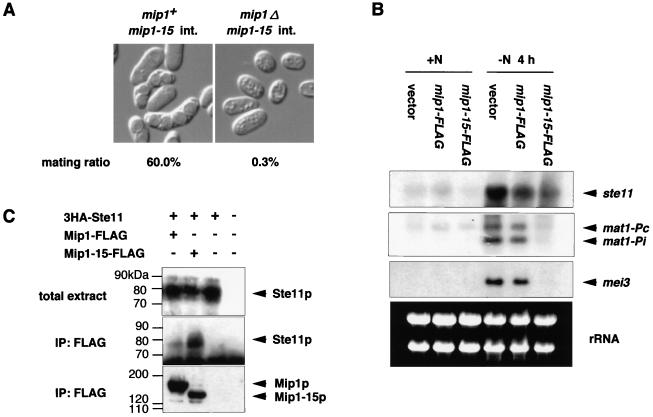
In fission yeast, the onset of meiosis is triggered by activation of the RNA-binding protein Mei2p. We screened for a high-copy-number suppressor of the ectopic meiosis induced by expression of an active form of Mei2p. Consequently we isolated a truncated form of a novel gene, named mip1, from a fission yeast genomic library. The mip1 gene encoded a protein of 1,313 amino acids which carried a WD-repeat motif in the C-terminal region and was apparently conserved among eukaryotes. Mip1p was cytoplasmic, and two-hybrid and immunoprecipitation analyses demonstrated that Mip1p was bound to Mei2p in vivo. Genetic evidence indicated that wild-type Mip1p was required for the function of Mei2p to induce meiosis and that the truncated form of it (Mip1-15p) dominantly interfered with Mei2p. Mip1p appeared to be involved also in conjugation, associating with Ste11p, which is a key transcription factor for sexual development. Furthermore, Mip1p was essential for cell growth, to which neither Mei2p nor Ste11p is relevant. These results suggest that Mip1p assists functional expression of a number of proteins required for proliferation and sexual development in fission yeast.
Figures

References
-
- Bai C, Elledge S J. Gene identification using the yeast two-hybrid system. Methods Enzymol. 1996;273:331–347. - PubMed
-
- Beach D, Rodgers L, Gould J. RAN1+ controls the transition from mitotic division to meiosis in fission yeast. Curr Genet. 1985;10:297–311. - PubMed
-
- Bresch C, Muller G, Egel R. Genes involved in meiosis and sporulation of a yeast. Mol Gen Genet. 1968;102:301–306. - PubMed
-
- Egel R. Mating-type genes, meiosis, and sporulation. In: Nasim A, et al., editors. Molecular biology of the fission yeast. San Diego, Calif: Academic Press; 1989. pp. 31–73.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases