

Decoding temporal information: A model based on short-term synaptic plasticity
- PMID: 10648718
- PMCID: PMC6774169
- DOI: 10.1523/JNEUROSCI.20-03-01129.2000
Decoding temporal information: A model based on short-term synaptic plasticity
Abstract
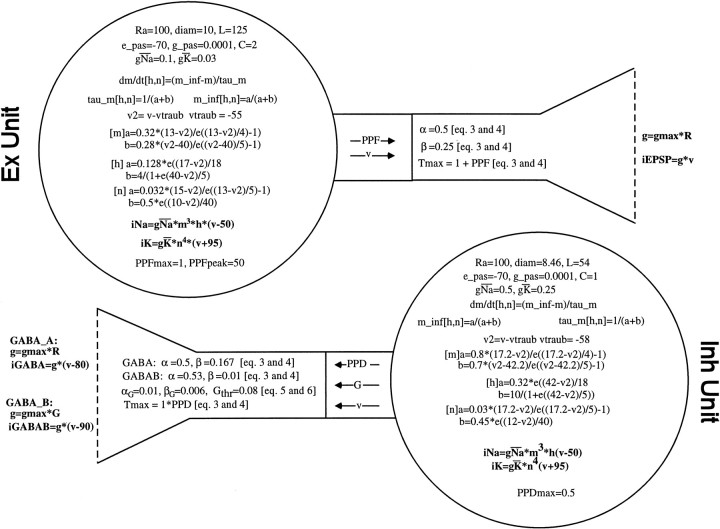
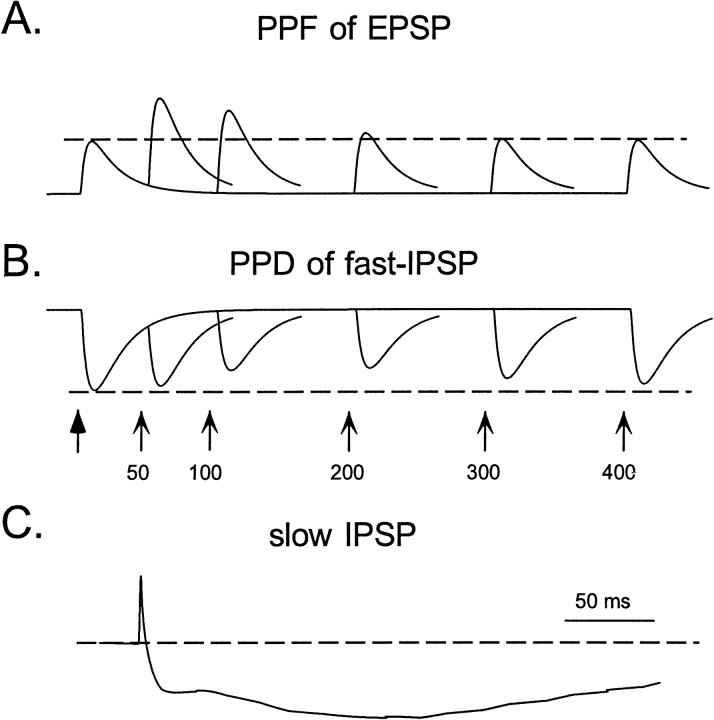
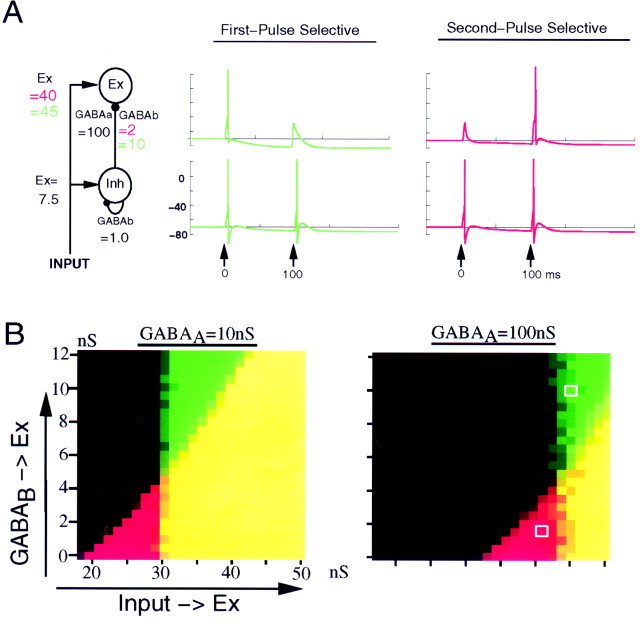
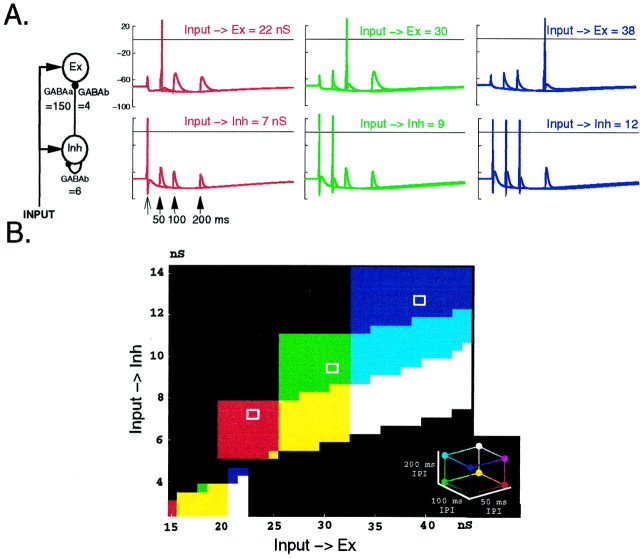
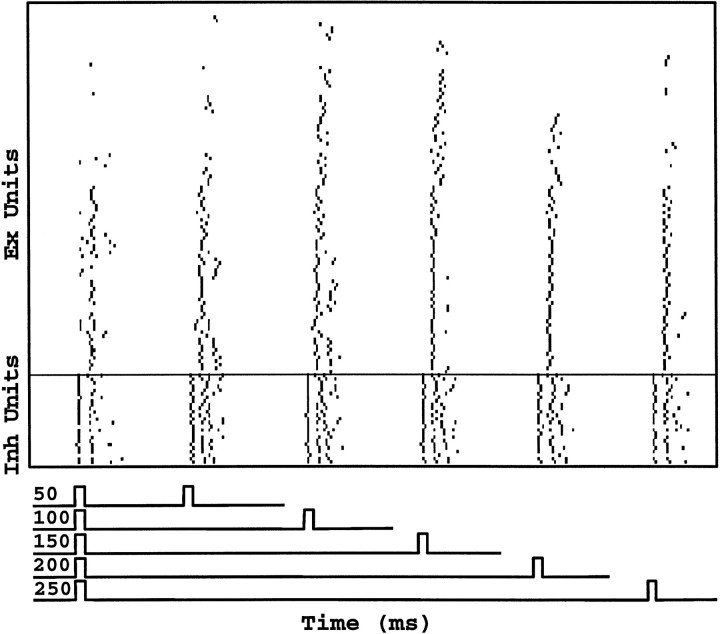
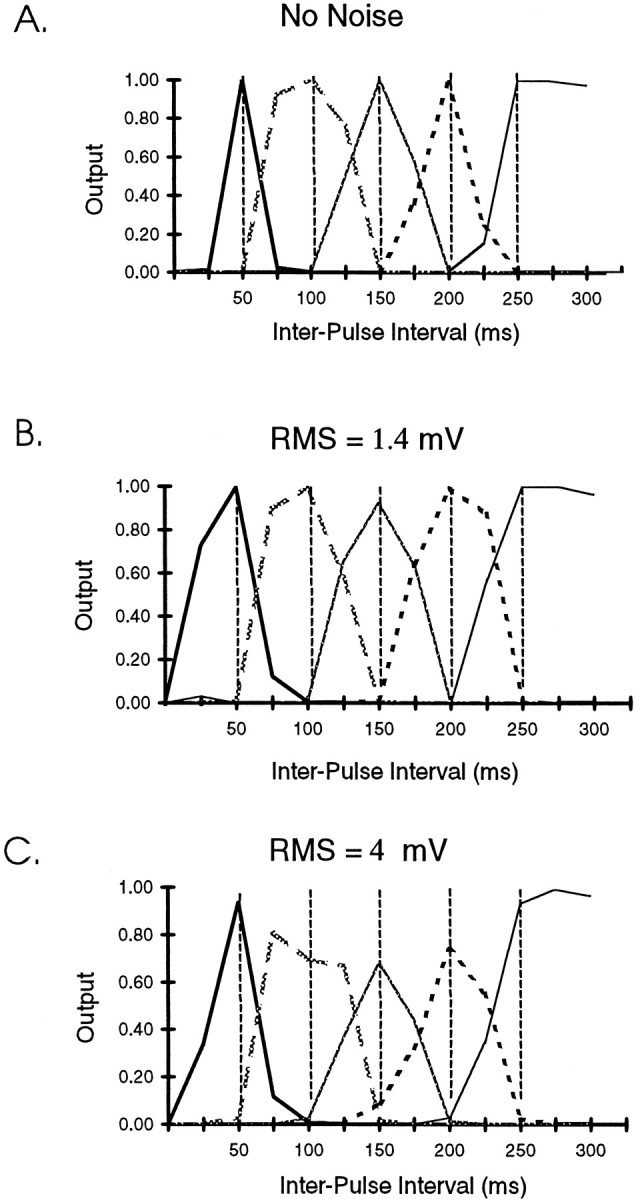
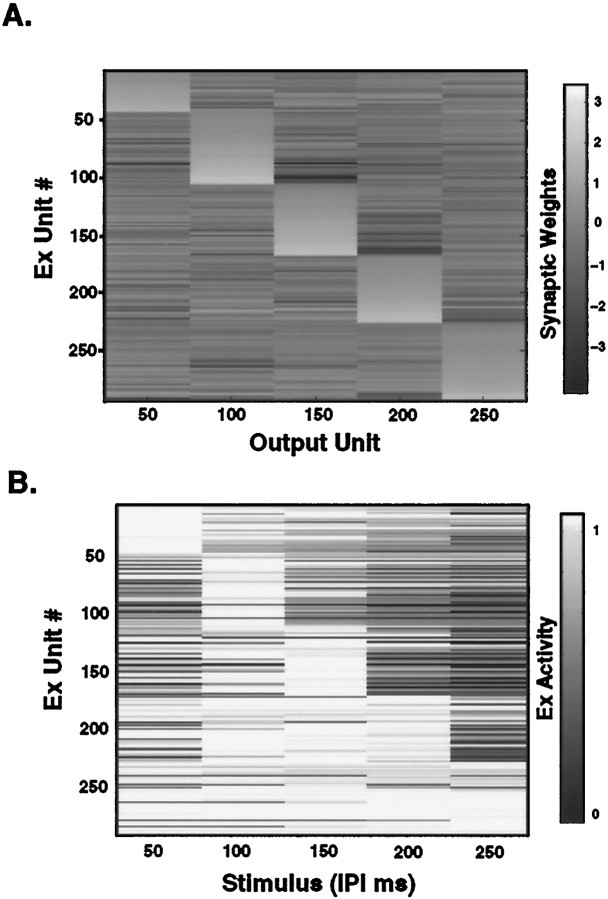
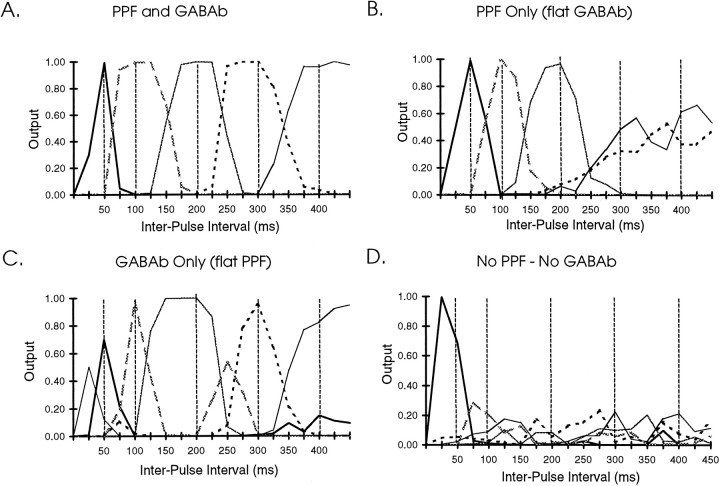
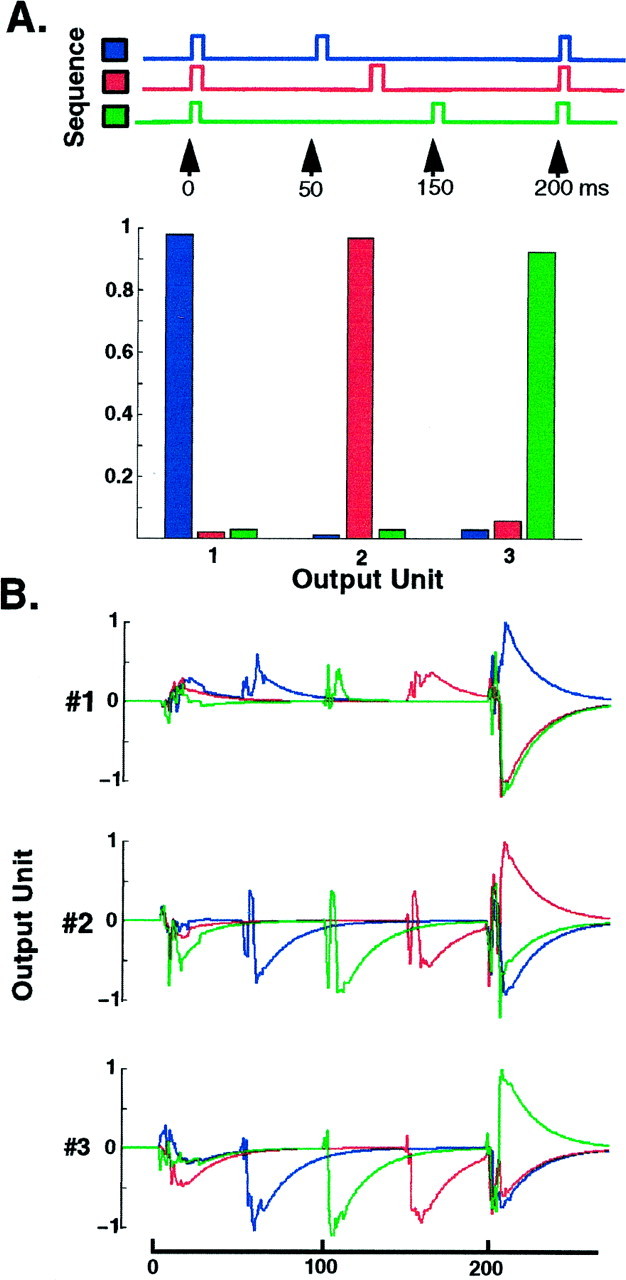
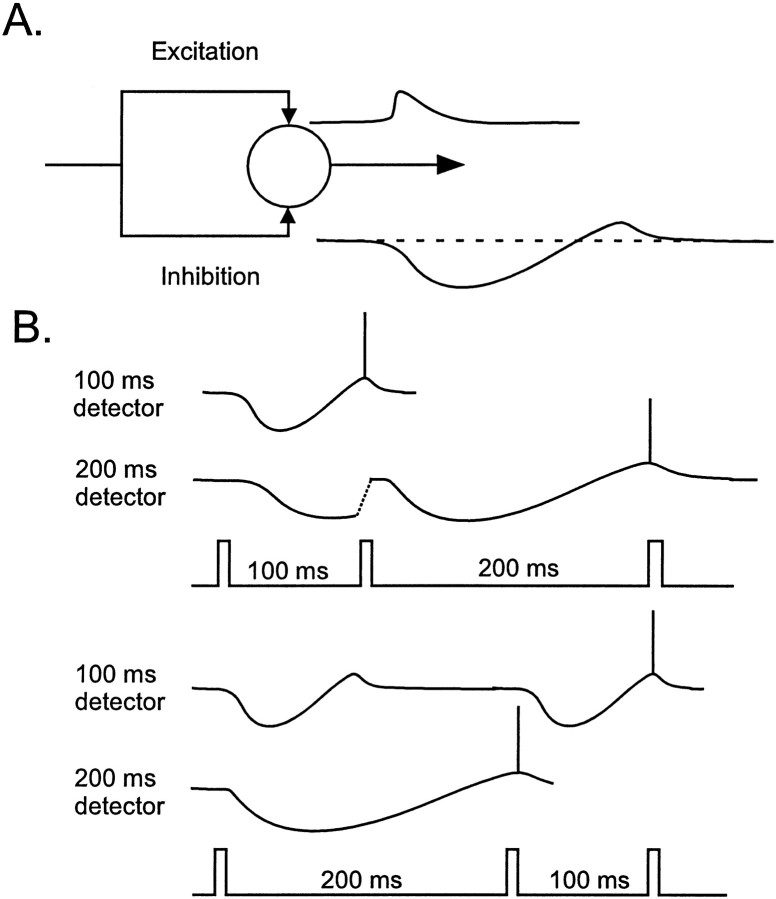
In the current paper it is proposed that short-term plasticity and dynamic changes in the balance of excitatory-inhibitory interactions may underlie the decoding of temporal information, that is, the generation of temporally selective neurons. Our initial approach was to simulate excitatory-inhibitory disynaptic circuits. Such circuits were composed of a single excitatory and inhibitory neuron and incorporated short-term plasticity of EPSPs and IPSPs and slow IPSPs. We first showed that it is possible to tune cells to respond selectively to different intervals by changing the synaptic weights of different synapses in parallel. In other words, temporal tuning can rely on long-term changes in synaptic strength and does not require changes in the time constants of the temporal properties. When the units studied in disynaptic circuits were incorporated into a larger single-layer network, the units exhibited a broad range of temporal selectivity ranging from no interval tuning to interval-selective tuning. The variability in temporal tuning relied on the variability of synaptic strengths. The network as a whole contained a robust population code for a wide range of intervals. Importantly, the same network was able to discriminate simple temporal sequences. These results argue that neural circuits are intrinsically able to process temporal information on the time scale of tens to hundreds of milliseconds and that specialized mechanisms, such as delay lines or oscillators, may not be necessary.
Figures
References
-
- Abbott LF, Varela JA, Kamal S, Nelson SB. Synaptic depression and cortical gain control. Science. 1997;275:220–224. - PubMed
-
- Adler TB, Rose GJ. Long-term temporal integration in the anuran auditory system. Nat Neurosci. 1998;1:519–523. - PubMed
-
- Beaulieu C, Kisvarday Z, Somogyi P, Cynader M, Cowey A. Quantitative distribution of GABA-immunopositive and -immunonegative neurons and synapses in the monkey striate cortex (area 17). Cereb Cortex. 1992;2:295–309. - PubMed
-
- Braitenberg V. Is the cerebellar cortex a biological clock in the millisecond range? Prog Brain Res. 1967;25:334–336. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources