

Rhomboid and Star facilitate presentation and processing of the Drosophila TGF-alpha homolog Spitz
- PMID: 10652272
- PMCID: PMC316351
Rhomboid and Star facilitate presentation and processing of the Drosophila TGF-alpha homolog Spitz
Abstract
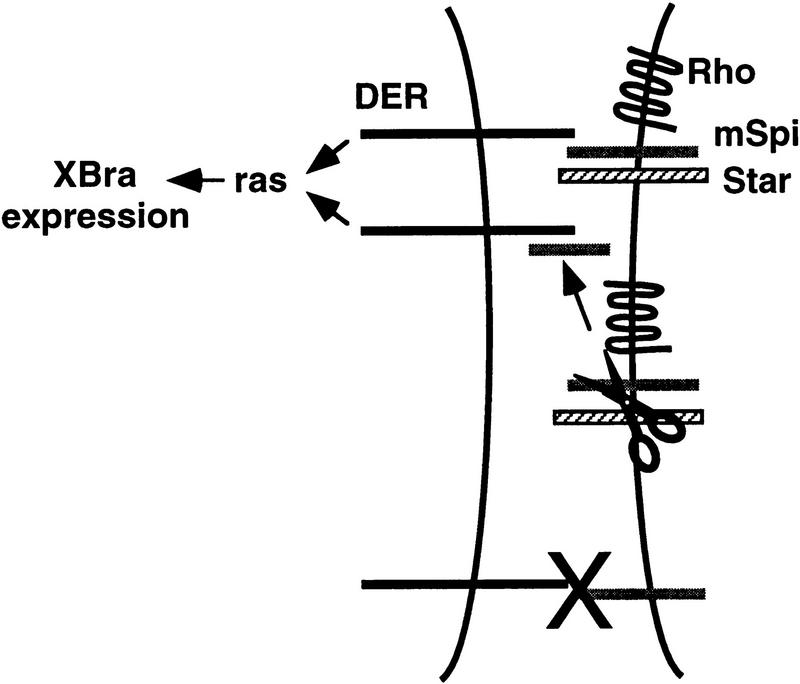
Activation of the Drosophila epidermal growth factor receptor (DER) by the transmembrane ligand, Spitz (Spi), requires two additional transmembrane proteins, Rhomboid and Star. Genetic evidence suggests that Rhomboid and Star facilitate DER signaling by processing membrane-bound Spi (mSpi) to an active, soluble form. To test this model, we use an assay based on Xenopus animal cap explants in which Spi activation of DER is Rhomboid and Star dependent. We show that Spi is on the cell surface but is kept in an inactive state by its cytoplasmic and transmembrane domains; Rhomboid and Star relieve this inhibition, allowing Spi to signal. We show further that Spi is likely to be cleaved within its transmembrane domain. However, a mutant form of mSpi that is not cleaved still signals to DER in a Rhomboid and Star-dependent manner. These results suggest strongly that Rhomboid and Star act primarily to present an active form of Spi to DER, leading secondarily to the processing of Spi into a secreted form.
Figures
Similar articles
-
Secreted Spitz triggers the DER signaling pathway and is a limiting component in embryonic ventral ectoderm determination.Genes Dev. 1995 Jun 15;9(12):1518-29. doi: 10.1101/gad.9.12.1518. Genes Dev. 1995. PMID: 7601354
-
Intracellular trafficking by Star regulates cleavage of the Drosophila EGF receptor ligand Spitz.Genes Dev. 2002 Jan 15;16(2):222-34. doi: 10.1101/gad.214202. Genes Dev. 2002. PMID: 11799065 Free PMC article.
-
brother of rhomboid, a rhomboid-related gene expressed during early Drosophila oogenesis, promotes EGF-R/MAPK signaling.Dev Biol. 2000 Oct 15;226(2):255-66. doi: 10.1006/dbio.2000.9851. Dev Biol. 2000. PMID: 11023685
-
EGF receptor signalling: the importance of presentation.Curr Biol. 2000 May 18;10(10):R388-91. doi: 10.1016/s0960-9822(00)00485-1. Curr Biol. 2000. PMID: 10837218 Review.
-
Twin peaks: Spitz and Argos star in patterning of the Drosophila egg.Cell. 1998 Oct 30;95(3):291-4. doi: 10.1016/s0092-8674(00)81759-6. Cell. 1998. PMID: 9814698 Review. No abstract available.
Cited by
-
A family of Rhomboid intramembrane proteases activates all Drosophila membrane-tethered EGF ligands.EMBO J. 2002 Aug 15;21(16):4277-86. doi: 10.1093/emboj/cdf434. EMBO J. 2002. PMID: 12169630 Free PMC article.
-
Expression in mammalian cell cultures reveals interdependent, but distinct, functions for Star and Rhomboid proteins in the processing of the Drosophila transforming-growth-factor-alpha homologue Spitz.Biochem J. 2002 Apr 15;363(Pt 2):347-52. doi: 10.1042/0264-6021:3630347. Biochem J. 2002. PMID: 11931664 Free PMC article.
-
Identifying pathways modulating sleep duration: from genomics to transcriptomics.Sci Rep. 2017 Jul 4;7(1):4555. doi: 10.1038/s41598-017-04027-7. Sci Rep. 2017. PMID: 28676676 Free PMC article.
-
Long-range signal transmission in autocrine relays.Biophys J. 2003 Feb;84(2 Pt 1):883-96. doi: 10.1016/S0006-3495(03)74906-6. Biophys J. 2003. PMID: 12547771 Free PMC article.
-
Insect oenocytes: a model system for studying cell-fate specification by Hox genes.J Anat. 2001 Jul-Aug;199(Pt 1-2):25-33. doi: 10.1046/j.1469-7580.2001.19910025.x. J Anat. 2001. PMID: 11523826 Free PMC article. Review.
References
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
-
- Bier E, Jan LY, Jan YN. rhomboid, a gene required for dorsoventral axis establishment and peripheral nervous system development in Drosophila melanogaster. Genes & Dev. 1990;4:190–203. - PubMed
-
- Brachmann R, Lindquist PB, Nagashima M, Kohr W, Lipari T, Napier M, Derynck R. Transmembrane TGF-alpha precursors activate EGF/TGF-alpha receptors. Cell. 1989;56:691–700. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous