

Antiapoptotic activity of Stat5 required during terminal stages of myeloid differentiation
- PMID: 10652277
- PMCID: PMC316353
Antiapoptotic activity of Stat5 required during terminal stages of myeloid differentiation
Abstract
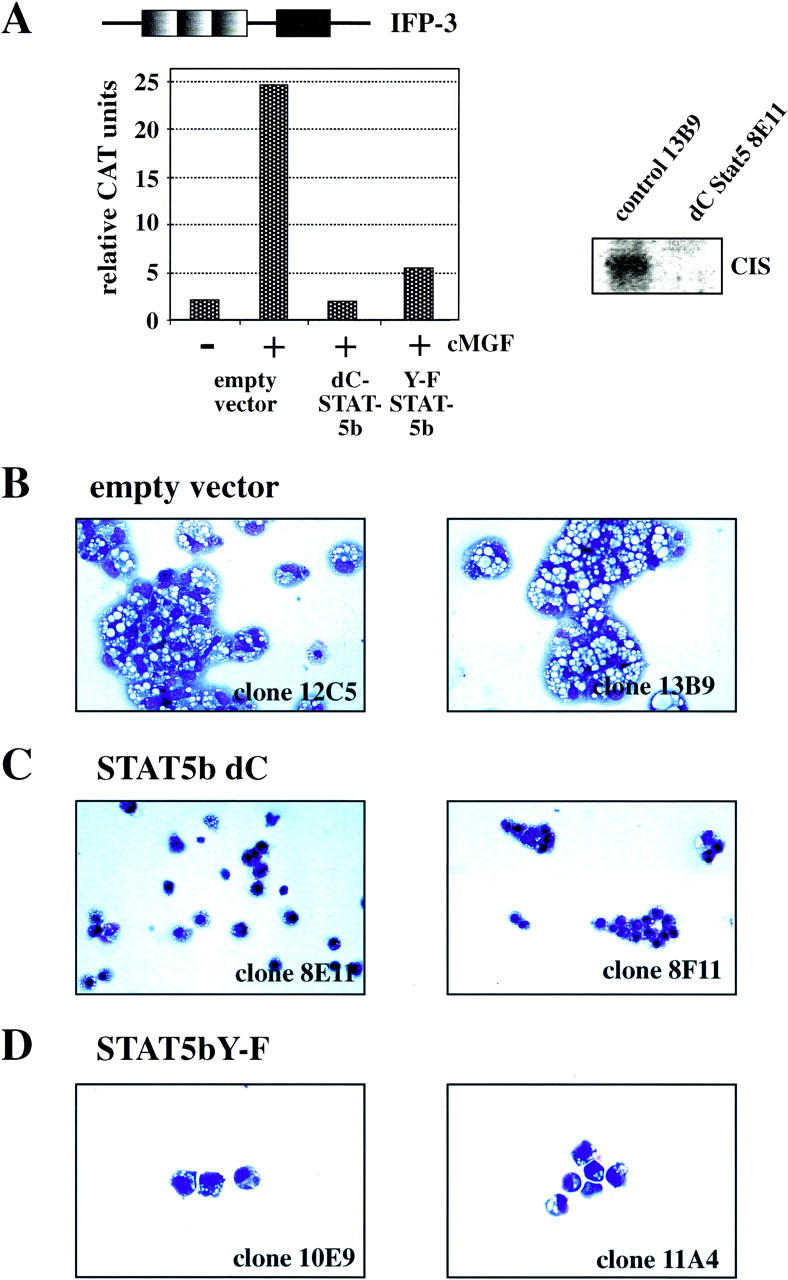
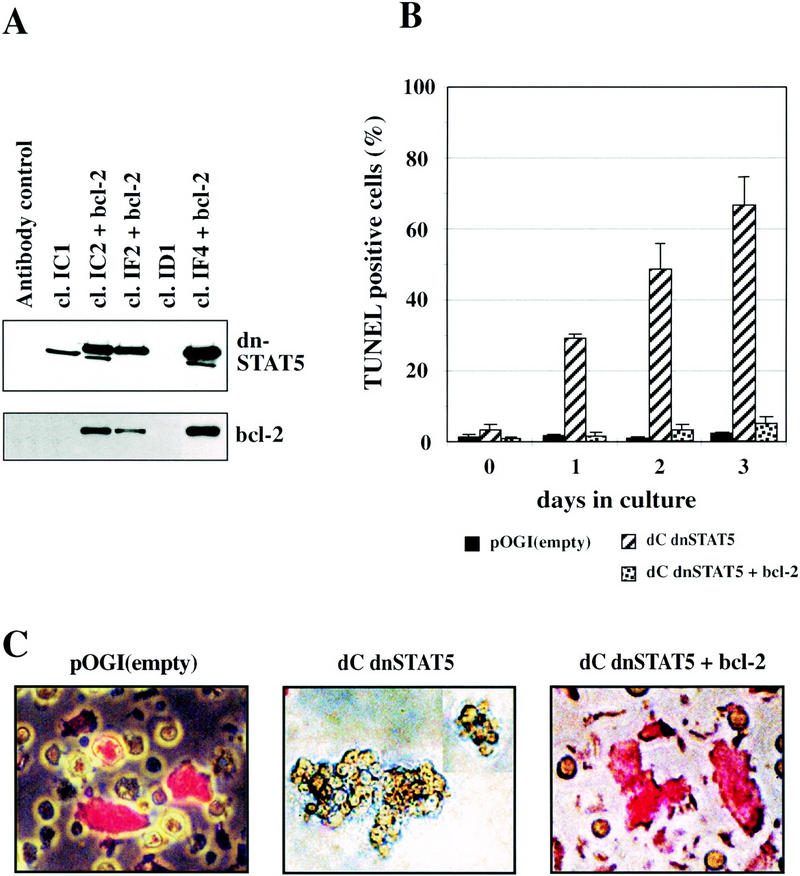
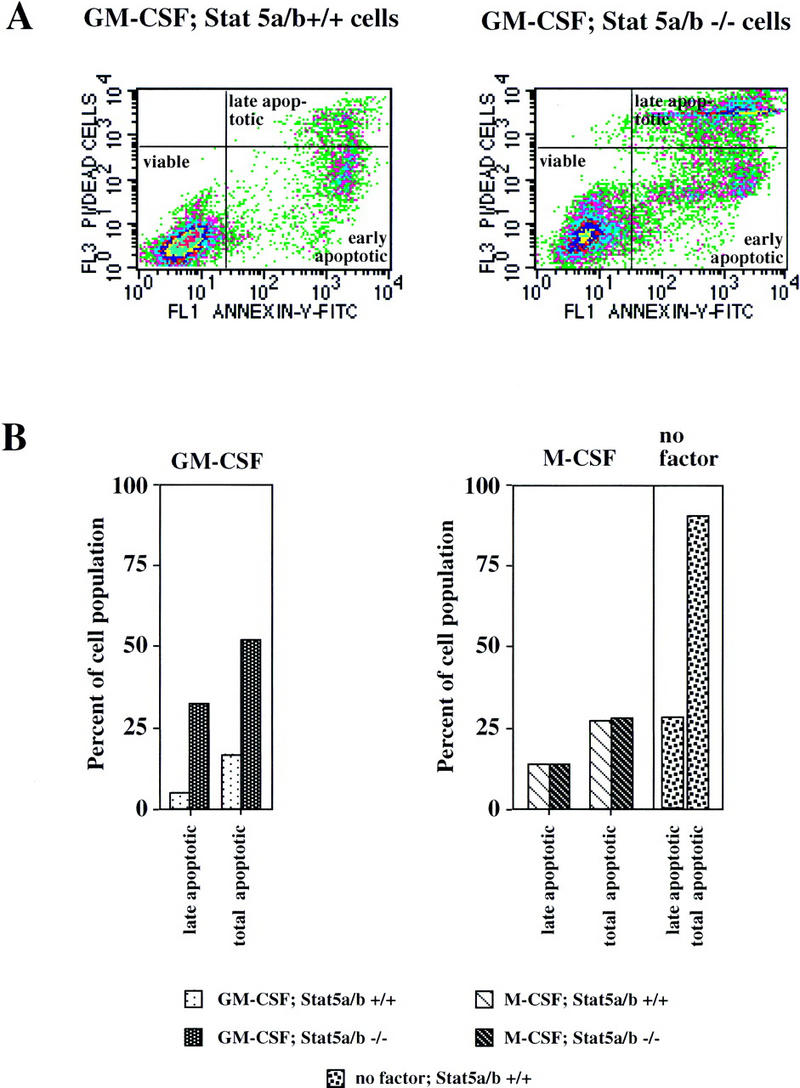
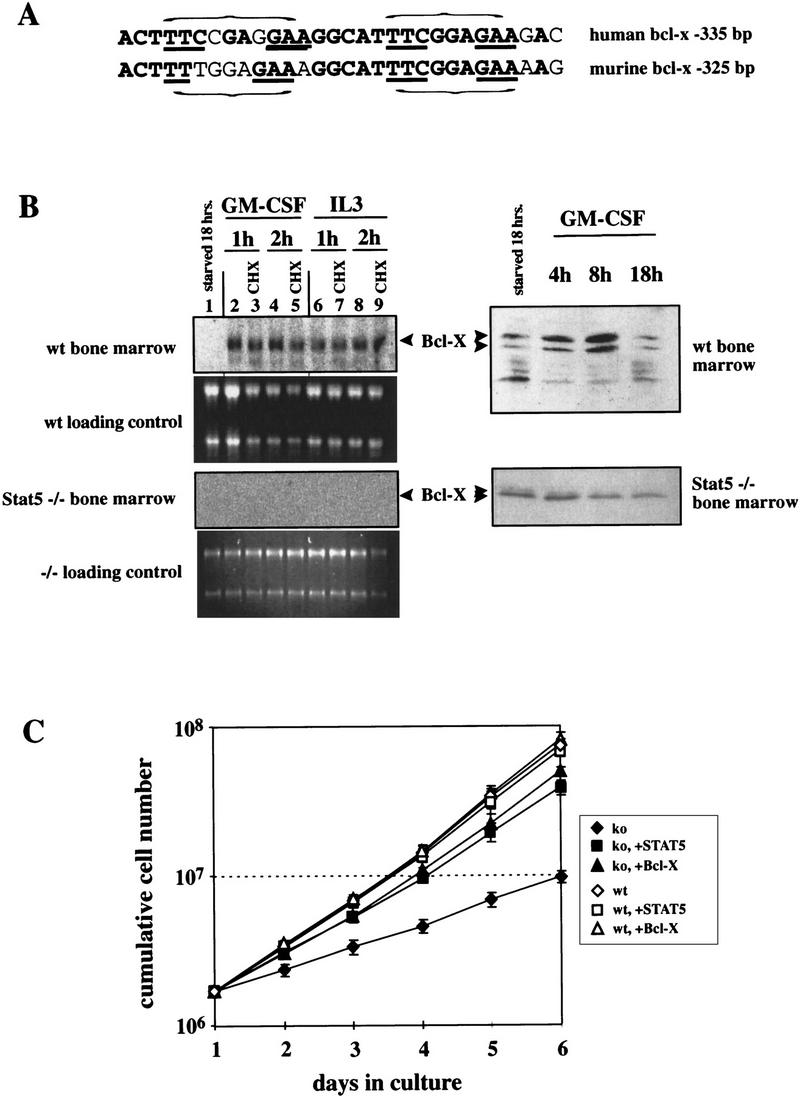
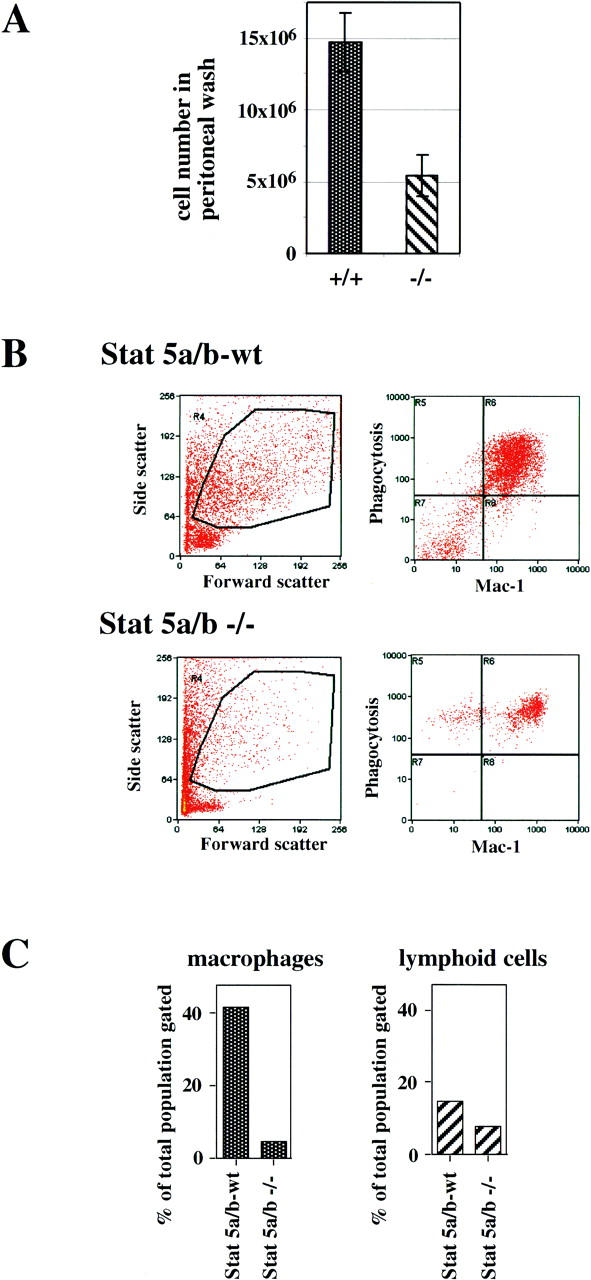
Stat5 is activated by multiple receptors of hematopoietic cytokines. To study its role during hematopoiesis, we have generated primary chicken myeloblasts expressing different dominant-negative (dn) alleles of Stat5. This caused a striking inability to generate mature cells, due to massive apoptosis during differentiation. Bcl-2 was able to rescue differentiating cells expressing dnStat5 from apoptosis, suggesting that during cytokine-dependent differentiation the main function of the protein is to ensure cell survival. Our findings with dnStat5-expressing chicken myeloblasts were confirmed with primary hematopoietic cells from Stat5a/Stat5b-deficient mice. Bone marrow cells from these animals displayed a strong increase in apoptotic cell death during GM-CSF-dependent functional maturation in vitro. The antiapoptotic protein Bcl-x was induced by GM-CSF and IL-3 in a Stat5-dependent fashion. Ectopic expression of Bcl-x rescued Stat5-deficient bone marrow cells from apoptosis, indicating that Stat5 promotes the survival of myeloid progenitor cells through its ability to induce transcription of the bcl-x gene. Finally, the recruitment of myeloid cells to inflammatory sites was found strongly impeded in Stat5-deficient mice. Taken together, our findings suggest that Stat5 may promote cytokine-dependent survival and proliferation of differentiating myeloid progenitor cells in stress or pathological situations, such as inflammation.
Figures


References
-
- Barahmand-pour F, Meinke A, Eilers A, Gouilleux F, Groner B, Decker T. Colony-stimulating factors and interferon-gamma activate a protein related to MGF-Stat 5 to cause formation of the differentiation-induced factors in myeloid cells. FEBS Letters. 1995;360:29–33. - PubMed
-
- Bedi A, Sharkis SJ. Mechanisms of cell commitment in myeloid cell differentiation. Curr Opin Hematol. 1995;2:12–21. - PubMed
-
- Beug H, Leutz A, Kahn P, Graf T. Ts mutants of E26 leukemia virus allow transformed myeloblasts, but not erythroblasts or fibroblasts, to differentiate at the nonpermissive temperature. Cell. 1984;39:579–588. - PubMed
-
- Beug H, Blundell PA, Graf T. Reversibility of differentiation and proliferative capacity in avian myelomonocytic cells transformed by tsE26 leukemia virus. Genes & Dev. 1987;1:277–286. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous